
HER – 2 / NEU STATUS EN ADJUVANTE ENDOCRIENE THERAPIE VOOR MAMMACARCINOOM

UNIVERSITEIT GENT
FACULTEIT DIERGENEESKUNDE
Academiejaar 2012- 2013
DE ROL VAN DE ENDOCRIENE PANCREAS IN HET KADER VAN PRODUCTIE EN GEZONDHEID
BIJ HOOGPRODUCTIEVE MELKKOEIEN
door
Hannes BOGAERT
Promotoren: Prof. dr. Geert Opsomer Literatuurstudie in het kader
Drs. Jenne De Koster van de Masterproef
©2013 Hannes Bogaert


Universiteit Gent, haar werknemers of studenten bieden geen enkele garantie met betrekking tot de juistheid of
volledigheid van de gegevens vervat in deze masterproef, noch dat de inhoud van deze masterproef geen inbreuk
uitmaakt op of aanleiding kan geven tot inbreuken op de rechten van derden.
Universiteit Gent, haar werknemers of studenten aanvaarden geen aansprakelijkheid of verantwoordelijkheid voor
enig gebruik dat door iemand anders wordt gemaakt van de inhoud van de masterproef, noch voor enig vertrouwen
dat wordt gesteld in een advies of informatie vervat in de masterproef.

UNIVERSITEIT GENT
FACULTEIT DIERGENEESKUNDE
Academiejaar 2012- 2013
DE ROL VAN DE ENDOCRIENE PANCREAS IN HET KADER VAN PRODUCTIE EN GEZONDHEID
BIJ HOOGPRODUCTIEVE MELKKOEIEN
door
Hannes BOGAERT
Promotoren: Prof. dr. Geert Opsomer Literatuurstudie in het kader
Drs. Jenne De Koster van de Masterproef
©2013 Hannes Bogaert

VOORWOORD
Hierbij zou ik graag alle personen willen bedanken die meegeholpen hebben aan het voltooien van deze
literatuurstudie.
In de eerste plaats wil ik Prof. dr. Opsomer bedanken. Eerst en vooral omdat hij mij de kans heeft
gegeven om over dit interessant onderwerp een literatuurstudie te schrijven. Daarnaast ook voor zijn vele
advies, zijn tijd die hij voor mij heeft vrijgemaakt om uitleg en informatie te geven en voor zijn
enthousiasme en aanmoediging.
Vervolgens wil ik ook mijn 2de
promotor Drs. Jenne De Koster bedanken voor zijn tijd die hij heeft
vrijgemaakt, ondanks hij het zelf zeer druk had, voor zijn uitleg, advies en hulp bij het schrijven.
Tot slot wil ik ook mijn ouders, broers en vrienden bedanken voor hun steun, hulp en aanmoediging
tijdens het schrijven van deze literatuurstudie.

INHOUDSPAGINA
AFKORTINGEN ............................................................................................................................................. 1
SAMENVATTING .......................................................................................................................................... 2
INLEIDING ..................................................................................................................................................... 3
LITERATUURSTUDIE ................................................................................................................................... 4
1. Melkproductie ........................................................................................................................................ 4
1.1. Algemene kenmerken ..................................................................................................................... 4
1.2. Fysiologie van de melkproductie ..................................................................................................... 4
1.2.1 De samenstelling van melk .................................................................................................... 4
1.2.2 De energiebalans bij het hoogproductief melkvee. ............................................................... 5
1.2.3 De lactatiecurve, met de link naar de energiebalans bij hoogproductief melkvee .............. 10
2. Gezondheid-metabole ziekten bij hoogproductief melkvee ................................................................. 14
2.1. Inleiding ......................................................................................................................................... 14
2.2. Bespreking ziekten ........................................................................................................................ 15
2.2.1 Fat cow syndrome = leververvetting = hepatic lipidosis = fatty liver en ketonemie ............. 15
2.2.2 Lebmaagverplaatsing .......................................................................................................... 19
2.2.3 Andere metabole ziekten ..................................................................................................... 20
3. De pancreas ........................................................................................................................................ 21
3.1. Anatomie ....................................................................................................................................... 21
3.2. Rol van de endocriene pancreas .................................................................................................. 22
3.2.1. Inleiding ............................................................................................................................... 22
3.2.2. Insuline ................................................................................................................................ 22
Algemeen .......................................................................................................................................... 22
Rol in de productie ............................................................................................................................ 24
Rol in de gezondheid ........................................................................................................................ 26
Insuline resistentie en secretie ......................................................................................................... 27
3.2.3. Glucagon ............................................................................................................................. 28
3.2.4. Pancreas polypeptide .......................................................................................................... 28
3.2.5. Somatostatine Pancreas polypeptide .................................................................................. 29
3.2.6. Insuline amyloid polypeptide (IAPP) of amylin .................................................................... 29
BESPREKING ............................................................................................................................................. 30
REFERENTIELIJST .................................................................................................................................... 32

1
AFKORTINGEN
AcCoA Acetylco-enzymA
ACTH Adrenocorticotroop hormoon
APO Apoproteïnen
ATP Adenosine trifosfaat
BCS Body Conditie Score
BHB Beta-Hydroxyboterzuur
CPT1 Carnitine Palmitoyltransferase 1
DSO Droge Stof Opname
GH Groeihormoon
GLUT Glucose transporters
HF Holstein Friesian
IAPP Insuline amyloïd polypeptide of amylin
IGF-1 Insulin-like growth factor 1
IL-1 Interleukine-1
IR Insuline resistentie
LDA Left Displacement Abomasum = linker lebmaagverplaatsing
LH Luteïniserend hormoon
MaCoA Malonylco-enzym A
NEB Negatieve Energie Balans
NEFA Non-esterified fatty acid
PP Pancreas polypeptide
p.p. Post partum
proIAPP Pro-islet amyloid polypeptide
PGF2-alfa Prostaglandine F2-alfa
SARA Sub-Acute Ruminal Acidosis
SRIF Somatotropine release inhibiting factor, synoniem voor somatostatine
TG Triglyceriden
TSH Thyroïd stimulerend hormoon
VLDL Very Low Density Lipoproteïnen

2
SAMENVATTING
Bij de huidige hoogproductieve melkkoeien ziet men een sterk toegenomen melkproductie, die gepaard
gaat met een verhoogde incidentie van metabole ziekten zoals leververvetting, ketonemie, en
hypocalcemie. Deze hebben in de meeste gevallen een negatieve invloed op de productie en het is dus
belangrijk om deze tot een minimum te beperken.
De link tussen de productie en de gezondheid is bij dit type dieren al uitgebreid onderzocht, maar het is
niet eenvoudig om deze volledig te doorgronden daar beiden een multifactorieel karakter hebben. De
endocriene pancreas staat in voor het lipiden-, proteïnen- en koolhydraatmetabolisme en speelt derhalve
een belangrijke rol in deze link. De hormonen die door de pancreas geproduceerd worden zijn insuline,
glucagon, pancreas polypeptide, somatostatine en insuline amyloïdpolypeptide. Van al deze hormonen is
insuline het meest onderzocht omdat het de belangrijkste functie heeft.
Bij herkauwers is er een groot verschil inzake het glucosemetabolisme in vergelijking met de
monogastrische dieren. Op de eerste plaats wordt door de aanwezigheid van het voormagensysteem de
glucosespiegel niet zozeer bepaald door absorptie van glucose uit het maagdarmstelsel, maar wordt
glucose merendeels aangemaakt via de gluconeogenese in de lever uitgaande van vluchtige vetzuren die
vrijkomen uit de pens. Daarnaast zijn de uier en de foetus vooral glucose afhankelijk, dit in tegenstelling
tot de perifere weefsels zoals vet- en spierweefsel die wel in staat zijn gebruik te maken van andere
energiebronnen waaronder vluchtige en vrije vetzuren, ketonen en bepaalde aminozuren. Om hoge
melkproducties te bereiken is het noodzakelijk om zoveel mogelijk glucose voor te behouden voor de uier,
welke aldaar zal worden omgezet tot lactose die op zijn beurt via osmose het melkvolume bepaalt.
Als adapatie ziet men bij het begin van de lactatie een gedaalde insulinesecretie, wat zorgt voor een
gedaalde plasma insulinespiegel, in combinatie met een perifere insulineresistentie (IR), die voor gevolg
heeft dat de werking van insuline sterk geremd wordt.
De perifere IR, t.h.v. de spieren en het vet, zorgt ervoor dat deze weefsels minder glucose kunnen
opnemen en de meerderheid van de glucose wordt gespaard voor de baarmoeder en de uier. Dit is
mogelijk door de verschillen in glucose transportermoleculen (GLUT), nl. insuline-onafhankelijke (o.a. type
1 en 3) t.h.v. de uier en de uterus en insuline-afhankelijke (type 4) t.h.v. het vet- en spierweefsel.
De verminderde insulinesecretie ontstaat door onderdrukking van de endocriene pancreas. Het exacte
onderliggende mechanisme hiervan is nog niet achterhaald, maar onderzoek heeft aangetoond dat o.a.
een chronisch gestegen non-esterified fatty acid (NEFA)-gehalte, via depressie van de β-cellen hiervoor
verantwoordelijk is. Dit fenomeen is al terug te vinden bij neonatale kalveren, wat doet vermoeden dat er
een (epi)genetische achtergrond is.
Door deze IR en gedaalde insulineconcentratie is de lipolyse van de vetreserves verhoogd, daar insuline
normaliter de lipolyse remt. Dit is enerzijds nuttig tijdens de toplactatie omdat de dieren dan niet in staat
zijn om voldoende energie op te nemen via de voeding, en daarom in een negatieve energiebalans
verkeren en beroep moeten doen op hun energiereserves opgeslagen in het vetweefsel. Dit laatste mag
echter niet uit de hand lopen aangezien de vrijgekomen NEFA’s kunnen zorgen voor het ontstaan van
metabole stoornissen door o.a. een daling in energie opname en een toegenomen IR, waardoor de dieren
in een neerwaartse spiraal terecht kunnen komen.
Sleutelwoorden: Endocriene pancreas – Insuline – Insuline resistentie – Hoogproductief melkvee –
Metabole ziekten
Key words: Endocrine pancreas – Insulin – Insulin resistance – High-yielding dairy cows –
Metabolic diseases

3
INLEIDING
De melkproductie van koeien is de afgelopen decennia door een doorgedreven selectie sterk
toegenomen, dit eist dan ook een grote adaptatie van het lichaam en het management (van Arendonk en
Liinamo, 2003; Pryce et al., 2004; Opsomer et al. 2006; Bossaert et al., 2008a; De Koster en Opsomer,
2012).
In combinatie met de verhoogde melkproductie is er een verhoogde incidentie van metabole stoornissen
zoals leververvetting, ketonemie, lebmaagverplaatsing en hypocalcemie. Deze vinden voornamelijk plaats
tijdens de toplactatie, gedurende de eerste weken na het afkalven. Dit komt doordat in deze periode zeer
extreme aanpassingen van het lichaam gevraagd worden en de dieren zich vaak in een negatieve
energiebalans (NEB) bevinden. Dit ontstaat doordat het dier onvoldoende energie kan opnemen om aan
de grote energievraag te voldoen. De ziekten zorgen voor een groot economisch verlies, door o.a. een
ongewenste productiedaling en verhoogde behandelingskosten, en daarom is het belangrijk om de
gezondheid van de koeien optimaal te houden. De koeien van tegenwoordig dienen als het ware als echte
topsporters behandeld en verzorgd te worden (Morrow, 1976; Veenhuizen et al., 1991; Goff en Horst,
1997; Jorritsma et al., 2003; Bobe et al., 2004; Grummer et al., 2004; Kokkonen, 2005; Leblanc et al.,
2005; Opsomer et al. 2006; Bossaert et al., 2008a en b; Cools, 2008a).
Er gebeurt al veel onderzoek naar het verband tussen de productie en de gezondheid. Door het
multifactorieel karakter van beiden is het echter niet eenvoudig om een exacte link hiertussen te
achterhalen. Zo speelt niet enkel de genetica, maar ook het management (o.a. voeding, melkafname,…)
een belangrijke rol. Ook zijn er verschillende orgaansystemen o.a. de lever, de spieren, de uier en het
vetdepot bij betrokken en is er een grote individuele variatie waar te nemen. De betrokkenheid van de
verschillende organen in combinatie met het multifactorieel karakter zorgt dan ook voor een complexe
zoektocht waar nog velen het hoofd op zullen breken (Rukkwamsuk et al., 1999a en b; Herdt, 2000;
Hayirli, 2006; Kerestes, 2009; Roche et al., 2009; Bossaert, 2010).
Bij de herkauwers is er een groot verschil met de monogastrische dieren inzake het glucose metabolisme.
Dit komt in de eerste plaats door hun voormagensysteem, dat ervoor zorgt dat de opgenomen
koolhydraten niet rechtstreeks in het bloed terecht komen, maar dat ze wel energie uit slecht verteerbare
voedingsstoffen zoals cellulose kunnen halen. De glucose bij de koe wordt vnl. via de gluconeogenese in
de lever aangemaakt. Ook ziet men dat de baarmoeder en de uier vrijwel uitsluiend aangewezen zijn op
glucose, dit in tegenstelling tot de perifere weefsels zoals het vet- en spierweefsel (Bell, 1995; Opsomer,
1995; Hayirli, 2006; Bossaert et al., 2008b; Schoenberg en Overton, 2010).
Het doel van deze literatuurstudie is om de link tussen de productie en gezondheid bij hoogproductieve
melkkoeien verder te ontrafelen door de rol van de endocriene pancreas te belichten. De endocriene
pancreas speelt namelijk een centrale rol in de productie en gezondheid door de sturing van het lipiden-,
het proteïnen- en het koolhydraatmetabolisme. Hierbij speelt voornamelijk de werking van insuline, nl. de
insuline secretie en de perifere insuline gevoeligheid een belangrijke rol (Kahn, 1978; Hove, 1978;
Brockman en Laarveld, 1986; Sano et al., 1991; Bell, 1995; Opsomer et al., 1999; Hayirli, 2006; Pires et
al., 2007; Kerestes, 2009; Bossaert, 2010).

4
LITERATUURSTUDIE
1. MELKPRODUCTIE
1.1. ALGEMENE KENMERKEN
De laatste jaren is er een sterke stijging van de melkproductie bij het melkvee in Vlaanderen
waargenomen. Zo is in vergelijking met 1950 de productie bijna verviervoudigd (van Arendonk en Liinamo,
2003; De Koster en Opsomer, 2012) en is een productie van 10 000 kg of meer per lactatie geen
uitzondering meer (Bossaert et al., 2008b). Deze stijging in melkproductie is het resultaat van een
doorgedreven genetische selectie en optimalisatie van het management en de voeding (van Arendonk en
Liinamo, 2003; Pryce et al., 2004; Opsomer et al. 2006; Bossaert et al., 2008a).
1.2. FYSIOLOGIE VAN DE MELKPRODUCTIE
1.2.1 De samenstelling van melk
Melk is een complex product dat samengesteld is uit lipiden (vetten, energiebron), koolhydraten (suikers,
energiebron), proteïnen (eiwitten, aminozuren bron), vitaminen, mineralen (elektrolyten bron) en water.
Tussen species is er een groot verschil in melksamenstelling. De melk van Holstein Friesian (HF) koeien
(de bron van de meeste melk in de Westerse wereld) heeft een gemiddelde samenstelling van 3.35%
eiwit, 4.25% vet ,4.6% lactose en een watergehalte van ongeveer 87% (Frandson et al., 2003;
Cunningham et al., 2007; Akers et al., 2008; Van Zeveren, 2012).
Het dieet en de fase van lactatie hebben ook een invloed op de samenstelling. Zo zorgt een dieet met een
hoog gehalte aan niet-vezelige koolhydraten, zoals zetmeel, voor een toename in het vetgehalte en een
dieet met een hoog eiwit gehalte promoot een lichte stijging van het eiwitgehalte, maar dit laatste effect is
kleiner dan dit van het vetgehalte. Het dieet heeft weinig invloed op het koolhydraten gehalte. Daarnaast
is ook te zien dat de percentages vet en eiwit relatief hoog zijn in de eerste weken na het kalven, in de
volgende drie tot vier maanden geleidelijk dalen en later in de lactatie deze terug zullen stijgen als de
totale melkproductie daalt (Frandson et al., 2003).
De meeste van de vetten in de melk zijn triglyceriden (TG). Deze zijn opgebouwd uit 3 vetzuren en een
glycerol molecule. Om deze vetzuren te kunnen produceren is de uier afhankelijk van acetaat en beta-
hydroxyboterzuur (BHB), die worden geproduceerd in de pens (Frandson et al., 2003; Cunningham et al.,
2007).
Lactose (of melksuiker) is het belangrijkste koolhydraat in de melk en is een disaccharide die
samengesteld is uit glucose en galactose. Lactose zorgt via osmose voor de aantrekking van water in de
uier en is hierdoor bepalend voor de hoeveelheid melk die er geproduceerd wordt. Aldus speelt glucose,
als precursor voor galactose en lactose, een zeer belangrijke rol in de melkproductie (Knight et al., 1994;
Zhao et al., 1996; Bell and Bauman, 1997; Bossaert et al., 2008b).
De belangrijkste melkproteïnen zijn de caseïnes (Frandson et al., 2003; Cunningham et al., 2007).
Daarnaast is er een hoog gehalte calcium aanwezig in de melk, ongeveer 1,3 g per liter. Door deze hoge
vraag aan calcium bij hoogproductieve koeien kan dit leiden tot hypocalcemie (melkziekte of kalfziekte),
maar dit kan grotendeels verholpen worden door een goed management (Akers et al., 2008).
Daar de hormonen van de endocriene pancreas instaan voor de regulatie van het metabolisme van
lipiden, koolhydraten en proteïnen, is dus een belangrijke rol weggelegd voor dit deel van de pancreas in
de productie. Deze rol zal verder besproken worden (Opsomer, 1995; Bossaert et al., 2008b).

5
1.2.2 De energiebalans bij het hoogproductief melkvee.
De energiebalans geeft het verschil weer tussen de energieopname en energiebehoefte/afgifte. De
energieopname wordt bepaald door de energie-inhoud van het voeder en door de “droge stof opname”
(DSO), terwijl de energiebehoefte/afgifte vnl. wordt bepaald door de opname van voedingsstoffen in de
uier en de drachtige baarmoeder (Opsomer, 1995; Janssens en Hesta, 2011).
Tijdens de toplactatie heeft het hoogproductief melkvee vaak te kampen met een enegatieve
energiebalans. Dit komt doordat energieopname lager is dan de grote energiebehoefte voor de
melkproductie en er aldus een energietekort ontstaat (Kokkonen, 2005; Bossaert et al., 2008b).
De energie-inhoud van een voeder wordt bepaald door de drie basisvoedingsstoffen nl. eiwitten, vetten
en koolhydraten (Silbernagl en Despopoulos, 2008). Dankzij hun speciaal spijsverteringstelsel, nl. het
voormagensysteem kunnen herkauwers gebruik maken van andere voedingsgrondstoffen dan niet-
herkauwers. Dit is voordelig omdat ze zo in staat zijn om uit een aantal slecht verteerbare vezelrijke
voedingsstoffen, zoals cellulose, toch nog energie te halen. De met het voeder opgenomen koolhydraten
worden bij herkauwers voor het grootste deel via microbiële fermentatie in de pens omgezet tot vluchtige
vetzuren. Deze doen dienst als belangrijkste energieleveranciers bij herkauwers. Bij monogastrische
dieren daarentegen is glucose, die voornamelijk via de darm wordt opgenomen, de belangrijkste
energieleverancier (Van Soest, 1994; Opsomer, 1995; Bell en Bauman, 1997; Frandson et al., 2003;
Hayirli, 2006; Cunningham et al., 2007; Bossaert et al., 2008b; Deprez, 2010; Janssens en Hesta, 2011).
In tegenstelling tot de monogastrische dieren passeert bij de herkauwers maar een kleine hoeveelheid
glucose de voormagen en wordt er weinig glucose rechtstreeks uit het rantsoen ter hoogte van de darm
geabsorbeerd. Hierdoor is de glucose spiegel in bloed bij herkauwers lager dan bij de niet-herkauwers.
Het merendeel van de glucose in de bloedbaan bij het rund wordt opgebouwd via gluconeogenese in de
lever en de nier, waarbij de bouwstenen afkomstig zijn van de pensfermentatie en spierafbraak (Young,
1976; Bell en Bauman, 1997; Frandson et al., 2003; Hayirli, 2006; Bossaert et al., 2008b).
Het is dus belangrijk dat er een goede penswerking is. De flora moet voldoende aangepast zijn om grote
hoeveelheden krachtvoer te kunnen verwerken en het epitheel moet voldoende ontwikkeld zijn om grote
hoeveelheden bouwstenen, zoals de vluchtige vetzuren, te kunnen absorberen. Er is ook een goede
leverwerking nodig om voldoende glucose te kunnen aanmaken via de gluconeogenese (Beever, 2006;
Bossaert et al., 2008b).
Bij het hoogproductief melkvee is er nog een extra wijziging inzake het koolhydraten metabolisme. De
uterus (groeiende foetus) en de uier zijn bijna uitsluitend aangewezen op glucose als energiebron en
kunnen amper overschakelen op andere energiebronnen. Dit in tegenstelling tot de perifere weefsels
zoals vetweefsel en spieren die wel vlot kunnen overschakelen op andere energiebronnen. Om aan deze
behoefte voor de uier en de baarmoeder te voldoen vindt er t.h.v. de perifere weefsel insuline resistentie
(IR) plaats en wordt m.a.w. de glucose opname zoveel mogelijk beperkt. Dit ‘glucose sparend’ effect zorgt
voor een voorkeur van nutriënten voor de dracht en melkproductie. Ook zal de gluconeogenese t.h.v. de
lever toenemen om voldoende glucose aan te kunnen maken (Bell, 1995; Opsomer, 1995; Hayirli, 2006;
Bossaert et al., 2008b; Schoenberg en Overton, 2010). Ter hoogte van de perifere weefsels zijn het
voornamelijk de vluchtige vetzuren, de vrije vetzuren (=Non-esterified fatty acid’s= NEFA’s), glycerol, de
ketonen en bepaalde aminozuren die de functie van glucose als energieleverancier overnemen (Herdt,
1988; Van Der Walt en Linington, 1989; Bell, 1995; Opsomer, 1995; Kokkonen, 2005; Bossaert et al.,
2008b).

6
Deze zullen we kort bespreken:
-Vluchtige vetzuren:
Zoals reeds eerder vermeld ontstaan door fermentatie van (plantaardige) koolhydraten in de pens
vluchtige vetzuren, vnl. azijnzuur (acetaat, C2), propionzuur (propionaat, C3), boterzuur (butyraat, C4) en
valeriaanzuur (C5) (Van Soest, 1994; Hayirli, 2006; Bossaert et al., 2008b; Janssens en Hesta, 2011).
Acetaat ontstaat vnl. wanneer er ruwvoeder (cellulose afbraak) wordt verstrekt. Daarnaast blijkt de lever
ook in staat acetaat te produceren, dit is vnl. tijdens de NEB. Gedurende deze periode wordt de lipolyse in
sterke mate gestimuleerd en worden er t.h.v. de lever veel vrije vetzuren afgebroken, wat op zijn beurt
aanleiding geeft tot een verhoogde release van acetaat. Dit acetaat, dat via de bloedbaan naar de perifere
weefsels wordt getransporteerd, blijkt een belangrijke energiebron te zijn (Van der Walt en Linington,
1989; Opsomer, 1995).
Propionaat wordt vnl. geproduceerd wanneer krachtvoerrijke (zetmeelafbraak) rantsoenen verstrekt
worden. Propionzuur is de belangrijkste glucogene bouwsteen voor de gluconeogenese. Via de portale
circulatie komt het in de lever terecht waar het gebruikt wordt voor de gluconeogenese (Frandson et al.,
2003; Bossaert et al, 2008b). Een klein deel komt niet in de lever terecht en kan dienst doen als
energiebron voor de perifere weefsels (Opsomer, 1995).
-Vetten:
Het vetweefsel dat vnl. uit TG bestaat is een van de belangrijkste energieopslagplaatsen waarin er steeds
opbouw en afbraak is. De balans is afhankelijk van de energetische status van het dier. Tijdens de NEB,
die vaak plaats vindt tijdens de eerste weken van de lactatie door de grote energievraag voor de
melkproductie en een gedaalde DSO, neemt de vetafbraak of lipolyse de bovenhand om aan de
energievraag te voldoen en worden de TG gehydrolyseerd met de vorming van 1 molecule glycerol en 3
moleculen NEFA’s die vrij komen in het bloed (Herdt, 2000; Rukkwamsuk et al., 2000; Bossaert et al.,
2008b).
De fysiologische en pathologische rol van deze NEFA’s is goed bestudeerd (De Koster en Opsomer,
2012). Ze worden verwerkt in de lever en dit gebeurt via drie verschillende wegen nl.volledige oxidatie,
gedeeltelijke oxidatie en re-esterificatie (Bell and Bauman, 1997; Rukkwamsuk et al., 1999a; Drackley et
al., 2005; Bossaert et al., 2008a; De Koster en Opsomer, 2012). In de levercel worden ze eerst omgezet
tot acylco-enzym A, vervolgens worden ze via de carnitine-shuttle opgenomen in de mitochondria waar ze
omgezet worden tot acetylco-enzymA (AcCoA) via de β-oxidatie en finaal wordt de AcCoA verwerkt in de
krebscyclus of de ketogenese. Vooral de opname in de mitochondria via de canitine shuttle en finale
verwerking zijn sterk afhankelijk van de energiestatus (Bossaert et al., 2008b). Tijdens energieoverschot
worden de NEFA’s niet opgenomen in de mitochondria, dit door een lage activiteit van het carnitine
palmitoyltransferase 1 (CPT1) door inhibitie door een hoge cytoplasmatische concentratie aan malonylco-
enzym A (MaCoA). De NEFA’s versmelten dan in het cytoplasma terug met glycerol met de vorming van
TG (re-esterificatie). Vervolgens vormen ze “Very Low Density Lipoproteïnen” (VLDL), door te versmelten
met apoproteïnen (APO), om dan verdeeld te worden over het lichaam (zie figuur 1) (Kräft, 2004; Bossaert
et al., 2008b; Bossaert, 2010). Tijdens energietekort is er een zeer intense lipolyse waardoor er een
overmaat aan NEFA’s ontstaat. Deze dringen wel de mitochondriën binnen, waar ze dan omgezet worden
in AcCoA. De verdere verwerking is dan opnieuw afhankelijk van de energiestatus. In energierijke
omstandigheden wordt het AcCoA, via de Krebs-cyclus finaal omgezet in CO2 en water met vrijkomen van
energie onder de vorm van adenosinetrifosfaat (ATP) (volledige oxidatie). Tijdens een NEB wordt

7
oxaloacetaat, onderdeel van de Krebs-cyclus, voornamelijk voorbehouden voor de gluconeogenese en
kan het AcCoA niet meer verwerkt in de Krebs-cyclus waardoor er een overschot aan AcCoA ontstaat.
Deze moleculen zullen met elkaar reageren tot de vorming van acetoacetaat, de moedermolecule van de
ketolichamen (onvolledige oxidatie met vorming van ketonen) (zie figuur 2) (Herdt, 1988; Bertics et
al.,1992; Opsomer, 1995; Goff en Horst,1997; Herdt, 2000; Bossaert et al., 2008b; Bossaert, 2010).
Zolang de vrijstelling van NEFA’s uit het vetweefsel beperkt is kan dit als nuttig aanzien worden aangezien
deze direct of indirect (als ketonen) gebruikt kunnen worden als energie substraat in verschillende
weefsels. Hierdoor kan er glucose gespaard worden voor de melkproductie en de baarmoeder. Echter een
zeer grote vetmobilisatie resulteert in een grote overvloed aan NEFA’s die een negatieve impact hebben
op de productie, reproductie, immuunstelsel en insuline werking. Zo is er een verminderde DSO, een
toegenomen IR en een toegenomen risico op leververvetting, ketonemenie, hypocalemie, retentio
secundinarium en mastitis (Holtenius and Holtenius 1996; Goff and Horst, 1997; Rukkwamsuk et al.,
2000; Bobe et al., 2004; Leroy et al., 2005; Pires et at., 2007; Bossaert et al., 2008c; Schoenberg en
Overton, 2010; De Koster en Opsomer, 2012).
Figuur 1. Tijdens energieoverschot is er een beperkte
lipolyse. De NEFA’s worden niet opgenomen in de
mitochondria in de levercel, dit door een lage activiteit
van carnitine palmitoyltransferase 1 (CPT1) door inhibitie
door een hoge cytoplasmatische concentratie aan
malonylco-enzym A (MaCoA). De NEFA’s versmelten in
het cytoplasma terug met glycerol met de vorming van
triacylglyceriden (TG). Vervolgens vormen ze “Very Low
Density Lipo-proteïnen” (VLDL) door te versmelten met
apoproteïnen (APO), om daarna terug verdeeld te
worden over het lichaam (uit Bossaert et al., 2008b;
Bossaert, 2010).
Figuur 2. Tijdens een energietekort is er zeer intense
lipolyse waardoor er een overmaat aan NEFA’s ontstaan.
De NEFA’s dringen de mitochondriën van de levercel
binnen, waar ze omgezet worden tot acetylco-enzymA
(AcCoA). Tijdens het tekort wordt oxalo-acetaat vnl.
voorbehouden voor de gluconeogenese en wordt het
AcCoA niet verwerkt in de Krebs-cyclus waardoor er een
overschot aan oxalo-acetaat ontstaat. Deze moleculen
zullen met elkaar reageren tot de vorming van
acetoacetaat, de moedermolecule van de ketonen. Bij
zeer intense lipolyse kunnen niet alle vetzuren naar de
mitochondria getransporteerd worden. Het teveel aan
vetten wordt opgeslagen als cytoplasmatische
vetglobulen. De metabolieten die in concentratie zijn
toegenomen zijn onderstreept (uit Bossaert et al., 2008b;
Bossaert, 2010).

8
Glycerol is een glucogene molecule die vooral in perioden van energietekort in beperkte mate kan
omgezet worden tot glucose (Bossaert et al., 2008b).
-Ketolichamen:
Ketolichamen kunnen ontstaan uit het voeder en zoals hoger vermeld ook ten tijde van de NEB, waarbij
de vele vrijgekomen NEFA’s worden omgezet tot ketonen. Dit laatste fenomeen staat bekend als de
ketogenese-gluconeogenese link (Herdt, 1988; Bertics et al., 1992; Opsomer, 1995; Goff en Horst, 1997;
Bossaert et al., 2008b).
De drie belangrijkste ketolichamen die bij het rund voorkomen zijn: aceto-acetaat, BHB en aceton
(Opsomer,1995; Deprez, 2010; Janssens en Hesta, 2011).
Boterzuur dat uit (vnl. slechte) kuilvoeder ontstaat, wordt t.h.v. de penswand omgezet tot BHB. Hierdoor
kan er vlug een hoge concentratie aan ketolichamen ontstaan wat aanleiding kan geven tot het ontstaan
van alimentaire ketonemie (Opsomer,1995).
Aceton komt in zeer kleine hoeveelheden voor in het plasma van de hoogproductieve koe en kan volgens
sommige auteurs in geringe mate omgezet worden in glucose (Herdt, 1988; Opsomer, 1995).
De ketolichamen kunnen in de perifere weefsels opgenomen worden en zorgen er voor een belangrijke
energiebijdrage (Kokkonen, 2005; Bossaert et al., 2008b).
-Aminozuren:
De aanvoer van propionzuur via de voeding is gelimiteerd waardoor er ook andere bronnen voor de
gluconeogenese nodig zijn tijdens de toplactatie. Hiervoor worden de spiereiwitten afgebroken en komen
er glucogene aminozuren (vnl. alanine en glutamine) vrij (Bossaert et al., 2008b).
De DSO is van groot belang bij het bepalen van de totale energieopname door het dier. Ze speelt een
belangrijke rol in de melkproductie en wordt aanzien als een belangrijke limiterende factor bij dieren die
genetisch in staat zijn tot zeer hoge producties (Bines, 1976; Bossaert et al., 2008b).
De voederopname bij melkvee wordt enerzijds bepaald door factoren die gekoppeld zijn aan het dier zelf,
zoals het ruimte innemende proces van de baarmoeder, stress rond de partus, de conditietoestand,
hormonen zoals leptine en insuline, metabolieten waaronder NEFA’s, puerperale stoornissen,
hypocalcemie, retentio secundinarum, endometritis, mastitis en kreupelheid. Anderzijds ook door factoren
die gekoppeld zijn aan het management, zoals het rantsoen, de voederingswijze (hoeveelheden,
frequentie van toedienen, samenstelling), de stalomgeving (bv. temperatuur, een hogere
omgevingstemperatuur zorgt voor een daling in de voederopname) en koppelgenoten (Nocek en Braund,
1985; Grummer et al., 2004; Beever, 2006; Bossaert et al., 2008a; Bossaert et al., 2008b). Daarbij dient
ook opgemerkt te worden dat de voederopnamecapaciteit van de huidige koeien sterk genetisch bepaald
is. Zo is het gestalte en buikvolume van de koeien de laatste decennia sterk toegenomen, wat de
capaciteit ten goede komt (Bossaert et al., 2008b). Ook zou dit verklaard kunnen worden doordat het
groter volume aan vetweefsel gevoeliger is voor glucocorticoïden en minder gevoelig is voor BHB, glucose
en insuline (Bobe et al., 2004; Drackley et al., 2005; Roche et al., 2009; De Koster en Opsomer, 2012).
De conditie of voedingstoestand bij het afkalven speelt een grote rol in de DSO. Bij koeien die te veel
vervetten aan het einde van de lactatie of tijdens de droogstand zal er een sterkere depressie in DSO
optreden, zal er een geringere toename van de totale opname na het afkalven zijn en is het tijdsinterval
tussen de maximale productie en de maximale voederopname toegenomen. Dit fenomeen is gebaseerd

9
op het feit dat zoogdieren een soort ingebouwd regelsysteem hebben meegekregen dat er voor zorgt dat
de voedselopname afneemt wanneer de lichaamsreserves toenemen. Hierbij speelt ook het NEFA-
gehalte een belangrijke rol, een verhoogd gehalte zorgt nl. voor een gedaalde inname. Anderzijds mogen
de reserves ook niet minimaal zijn aangezien zij een belangrijke energiebron zijn voor de melkproductie
gedurende de eerste maanden van de lactatie. Een reserve-tekort kan ervoor zorgen dat er geen snelle
stijging van de melkproductie is, de verwachte lactatiepiek niet bereikt wordt en de verwachte 305-
dagenproductie niet bereikt wordt (Garnsworthy en Topps, 1982; Wildman et al.,1982; Treacher et al.,
1986; Opsomer, 1995; Bossaert et al., 2008b; Bossaert, 2010; Schoenberg en Overton, 2010).
Bij toevoeging van een hoeveelheid krachtvoer aan het rantsoen, daalt de DSO van het ruwvoer
(verdringing van ruwvoer door krachtvoer). Wanneer het aandeel krachtvoeder in het rantsoen te groot
wordt (boven 70%) kunnen er problemen ontstaan met de penswerking, wat gepaard gaat met een
duidelijke daling van het melkvetgehalte (Opsomer, 1995; Cools et al., 2008b). Ook is er een daling van
de melkproductie en kunnen er metabole ziekten zoals leververvetting ontstaan (Morrow, 1976).
Voor en tijdens de kalving zijn er grote veranderingen te zien in de DSO, zo is er een daling van
voorbijgaande aard op te merken. De ernst van deze daling bepaalt in sterke mate ook de stijging na de
partus. Zo is geweten dat koeien die een hogere DSO opname hebben voor de partus, ook na de partus
een hogere opname hebben, en omgekeerd koeien die een grotere daling meemaken voor en rond de
partus bv. door vervetting tijdens de droogstand wat zorgt voor een sterkere toename van NEFA’s of
stress, een tragere stijging in DSO hebben en doorgaans ook langer in NEB blijven (Bertics et al., 1992;
Grummer et al., 2004; Bossaert et al., 2008a; Bossaert et al., 2008b). Na de kalving is de
energieaanvraag groot en zal een stijging in de DSO dit deels compenseren. Deze stijging loopt echter
niet gelijk met de stijgende energievraag voor de melkproductie in de vroege lactatie. Zo situeert de
lactatiepiek zich rond week 3 tot 7 post partum (p.p.) en zal de DSO zijn maximum pas bereiken tussen
week 8 en 22 p.p. waardoor de energie opname steeds achterloopt en de koe in een NEB zal verkeren.
Ter aanvulling zullen de energiereserves, vnl. vetten afgebroken worden waarbij dan de NEFA’s
vrijkomen. De NEB is nog meer uitgesproken bij de koeien die in een te hoge conditiescore afkalven
doordat er een verhoogde vrijstelling is van NEFA’s en er dus een ergere daling is in DSO. Hierdoor zullen
de vetreserves nog meer worden aangesproken waardoor de koeien dan in een negatieve spiraal terecht
komen (Grummer et al., 2004; Bossaert et al., 2008a; Bossaert et al., 2008b; Schoenberg en Overton,
2010).
Om deze reserves of m.a.w. de conditie, opgeslagen in vet of spierweefsel, in te schatten maakt men
gebruik van de body conditie score (BCS). Dit is een subjectieve methode waarbij de hoeveelheid vet- en
spierweefsel op specifieke plaatsen zo nauwkeurig mogelijk geschat wordt en een totaalscore wordt
bepaald. Vaak wordt gebruik gemaakt van het systeem van Edmondson et al. (1989) waarbij een score
wordt toegekend gaande van één (zeer mager) tot vijf (zeer vet) (zie figuur 3). Deze schaal kan verschillen
van land tot land (Roche et al., 2009).

10
Figuur 3. Scoringskaart voor het bepalen van de body condition score (BCS) (Edmonson et al., 1989)
1.2.3 De lactatiecurve, met de link naar de energiebalans bij hoogproductief melkvee
De hoge melkproductie bij melkvee vereist zeer veel energie. De energie die ze opnemen via het voeder
is niet voldoende om dit op te vangen en daarom moeten ze beroep doen op de reserves vnl. vetten die
ze ten tijde van energieoverschot hebben opgeslagen (Bossaert et al., 2008b). Doordat er perioden zijn
van energieoverschotten, waar er dan reserves worden aangelegd en perioden van energieschaarste,
waar de reserves worden aangesproken, en dit steeds opnieuw plaats vindt kan er gesproken worden van
een lactatiecyclus (zie figuur 4). Deze kan ingedeeld worden in 4 perioden namelijk een aanvang- of
vroege lactatie, midden lactatie, eindlactatie en de droogstand waarbij elke fase gekenmerkt is door zijn
eigen eigenschappen (Opsomer, 1995). Vaak wordt gebruikt gemaakt van de term transitieperiode. De
transitieperiode wordt beschreven als de periode van 3 weken voor en 3 weken na het afkalven en ze kan
onderverdeeld worden in 2 delen nl. de droogstand en het begin van de lactatie, waarbij de partus het
kantelmoment vormt (Bossaert et al., 2008b; Cools et al., 2008b; Hermans, 2010).
11
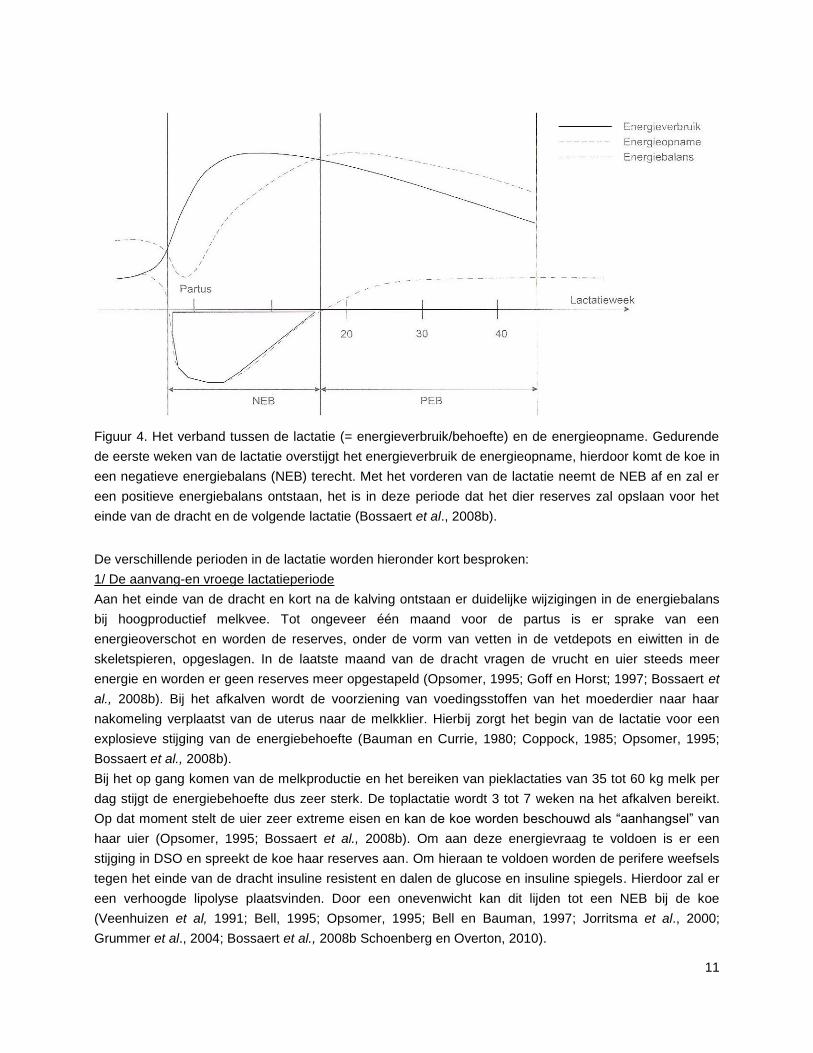
Figuur 4. Het verband tussen de lactatie (= energieverbruik/behoefte) en de energieopname. Gedurende
de eerste weken van de lactatie overstijgt het energieverbruik de energieopname, hierdoor komt de koe in
een negatieve energiebalans (NEB) terecht. Met het vorderen van de lactatie neemt de NEB af en zal er
een positieve energiebalans ontstaan, het is in deze periode dat het dier reserves zal opslaan voor het
einde van de dracht en de volgende lactatie (Bossaert et al., 2008b).
De verschillende perioden in de lactatie worden hieronder kort besproken:
1/ De aanvang-en vroege lactatieperiode
Aan het einde van de dracht en kort na de kalving ontstaan er duidelijke wijzigingen in de energiebalans
bij hoogproductief melkvee. Tot ongeveer één maand voor de partus is er sprake van een
energieoverschot en worden de reserves, onder de vorm van vetten in de vetdepots en eiwitten in de
skeletspieren, opgeslagen. In de laatste maand van de dracht vragen de vrucht en uier steeds meer
energie en worden er geen reserves meer opgestapeld (Opsomer, 1995; Goff en Horst; 1997; Bossaert et
al., 2008b). Bij het afkalven wordt de voorziening van voedingsstoffen van het moederdier naar haar
nakomeling verplaatst van de uterus naar de melkklier. Hierbij zorgt het begin van de lactatie voor een
explosieve stijging van de energiebehoefte (Bauman en Currie, 1980; Coppock, 1985; Opsomer, 1995;
Bossaert et al., 2008b).
Bij het op gang komen van de melkproductie en het bereiken van pieklactaties van 35 tot 60 kg melk per
dag stijgt de energiebehoefte dus zeer sterk. De toplactatie wordt 3 tot 7 weken na het afkalven bereikt.
Op dat moment stelt de uier zeer extreme eisen en kan de koe worden beschouwd als “aanhangsel” van
haar uier (Opsomer, 1995; Bossaert et al., 2008b). Om aan deze energievraag te voldoen is er een
stijging in DSO en spreekt de koe haar reserves aan. Om hieraan te voldoen worden de perifere weefsels
tegen het einde van de dracht insuline resistent en dalen de glucose en insuline spiegels. Hierdoor zal er
een verhoogde lipolyse plaatsvinden. Door een onevenwicht kan dit lijden tot een NEB bij de koe
(Veenhuizen et al, 1991; Bell, 1995; Opsomer, 1995; Bell en Bauman, 1997; Jorritsma et al., 2000;
Grummer et al., 2004; Bossaert et al., 2008b Schoenberg en Overton, 2010).

12
Daar in deze periode de lichaamsreserves gemobiliseerd worden is er een afname van het
lichaamsgewicht en een daling van de conditiescore, wat bij hoogproductief melkvee in deze periode als
“normaal” kan worden beschouwd (Opsomer, 1995; Bossaert et al., 2008a; Schoenberg en Overton,
2010). Gemiddeld situeert het diepste punt van de NEB zich rond 14 dagen p.p, waarbij er een grote
individuele variatie is tussen de koeien (Bossaert et al., 2008b; Bossaert, 2010).
2/ De midden- en laat-lactatie periode
Door de geleidelijke daling van de melkproductie en het stijgen van de voederopname, komt de
energiebalans rond de 10e tot 14
e week p.p. in evenwicht. Een verdere daling van de energiebehoefte en
een stijging van de energieopname brengt de dieren zelfs in een positieve energiebalans. In deze periode
zijn de dieren dan ook in staat de lichaamsreserves opnieuw aan te vullen (Opsomer 1995; Bossaert et
al., 2008b; Bossaert, 2010).
Wel is het zo dat het systeem waarbij lichaamsreserves worden betrokken in de melkproductie en het dier
zich gedurende de lactatie in een cyclus bevindt van gewichtsverlies en gewichtstoename, biologisch
inefficiënt is. Het produceren van melk uit energiereserve is minder efficiënt dan het rechtstreeks
produceren van melk uitgaande van de voeding. En de efficiëntie voor de melkproductie, uitgaande van
lichaamsreserves aangelegd tijdens de droogstand is nog lager. De voorbereiding van de volgende
lactatie begint dus al in de periode voor de droogstand (Moe, 1981; Opsomer,1995).
3/ De droogstand
De droogstand wordt gedefinieerd als de niet-melkgevende periode voorafgaand aan de daaropvolgende
lactatie. Dit is een zeer belangrijke periode voor melkkoeien, ze dient om het dier tot rust te laten komen
en zich voor te bereiden op de volgende lactatie (Hostens, 2008). Tijdens de droogstand dient de voeding,
huisvesting en het management van de dieren zo optimaal mogelijk te zijn en er op gericht dat de
‘topatleten’ zich op een ideale manier kunnen ‘opwarmen’ om aan hun ‘sprint’ te beginnen, nl. een
toplactatie en deze kunnen bereiken zonder dat zij worden aangetast door (metabole) stoornissen
(Opsomer, 1995; Cools et al., 2008b; Hostens, 2008; Schoenberg en Overton, 2010).
Bij de droogstand zijn er verschillende belangrijke parameters nl. namelijk de lengte van de droogstand,
het al dan niet werken met 2 verschillende groepen tijdens de droogstand en het rantsoen (Hostens,
2008).
De lengte van de droogstand
Over de lengte van de droogstand bestaat tot op heden heel wat discussie. Uit vroeger onderzoek blijkt
dat de melkproductie optimaal is wanneer de dieren na de intense lactatie gedurende een periode van 50
à 60 dagen of m.a.w. 7 tot 8 weken hebben drooggestaan (Coppock et al., 1974; Opsomer, 1995). In de
praktijk merken we dat deze veelal varieert van 0 tot 8 weken. Een verkorte droogstand periode bv. 4
weken tot volledig weglaten zou de volgende voordelen kunnen hebben: ten eerste, het wegvallen van de
noodzaak tot het voeren van een laag energetisch droogstandsrantsoen omdat de dieren bij een kortere
droogstand de kans niet meer krijgen om te vervetten en ten tweede, wordt de pens niet meer
geconfronteerd met abrupte rantsoenwisselingen die gehanteerd worden bij de overgangen lactatie-
droogstaand en omgekeerd. Hierbij is wel op te merken dat vaarzen meer behoefte hebben aan een
langere droogstand periode dan oudere koeien, daarom is dan ook een droogstandsperiode van 6 weken
voor vaarzen aan te raden, terwijl voor koeien een droogstandslengte van 5 weken kan volstaan (Hostens,
2008).

13
Werken met verschillende groepen
Het indelen van de droge koeien in twee groepen is een strategie die reeds langer geadviseerd wordt om
zo het management te kunnen optimaliseren nl. een “vroege droogstandsperiode” (early dry period of far-
off = periode net na het droogzetten) en “einde droogstandsperiode” (close-up dry period = periode net
voor het afkalven).
Het rantsoen
Bij het begin van de droogstand is er de energiebehoefte minimaal en bevinden de dieren zich in een
positieve energiebalans. Om dan een overmatige vervetting te voorkomenis het best dat het voeder niet te
energierijk is (Opsomer, 1995; Bossaert et al, 2008b; Cools et al., 2008b).
Tegen het einde van de droogstand (nl. groep “einde droogstandsperiode”) ontstaat er een geleidelijke
toename van de energiebehoeften, veroorzaakt door de grotere behoeften van de vrucht. Bij deze dieren
daalt de DSO, o.a. door het ruimte innemend proces van de uterus, zodat er energierijkere rantsoenen
moeten toegediend worden om dit deels te compenseren. Want als dieren afkalven in een te magere
conditie zullen ze, zoals hoger vermeld, hun genetische potentieel niet kunnen bereiken (Opsomer, 1995;
Bossaert et al., 2008b). Er wordt aangenomen dat een conditiescore van 3,5 rond het afkalven kan gelden
als algemeen streefdoel (Cools et al., 2008b).
Hoe goed het rantsoen ook in elkaar steekt, de DSO blijft de absolute nummer één voor wat betreft het
monitoren van het droogstandsmanagement. Het ten allen tijde op peil houden van de DSO rondom het
kalven blijft het ultieme doel (Opsomer, 1995; Bossaert et al., 2008b; Cools et al., 2008b).
Zoals reeds hoger vermeld is een goede penswerking onmisbaar voor een optimale lactatie. Na de kalving
is er een plotse overgang in voedersamenstelling en energievraag waarbij de pens zich op korte tijd moet
aanpassen aan een grotere DSO en grotere concentraties koolhydraatrijke voedingsstoffen. Deze fase
verloopt niet altijd probleemloos, zo kan een te bruuske overgang lijden tot verschillende problemen zoals
“subacute ruminal acidosis” (SARA), immunosuppressie, lebmaagverplaatsing en een verstoorde
voederopname en melkgift (Kleen et al., 2003; Beever, 2006; Bossaert et al., 2008b; Cools et al., 2008b).
Daarom wordt aanbevolen vanaf de twee laatste weken van de droogstand een kleine hoeveelheid
krachtvoer te verstrekken en de laatste twee weken voor de partus het lactatierantsoen aan te bieden
(Opsomer et al., 2004; Cools et al., 2008b).
Het risico op hypocalcemie of kalfziekte wordt eveneens grotendeels bepaald in de droogstand. Ter
preventie dient het droogstandsrantsoen te bestaan uit voedingsstoffen die arm zijn aan calcium en
kalium, zoals maïskuil en stro. Gras en graslandproducten voldoen niet aan deze vereisten en kunnen
daarom beter vermeden worden (Opsomer en de Kruif, 1998; Cools et al., 2008b; Hostens, 2008).

14
2. GEZONDHEID-METABOLE ZIEKTEN BIJ HOOGPRODUCTIEF MELKVEE
2.1. INLEIDING
De melkproductie is de laatste decennia sterk gestegen, de keerzijde hiervan is dat het optreden van
metabole stoornissen ook is toegenomen (van Arendonk en Liinamo, 2003; Bossaert et al., 2008a). Er zijn
veel metabole ziekten maar de vaakst voorkomende stoornissen zijn leververvetting, ketonemie,
hypocalcemie, lebmaagverplaatsing en pensverzuring (Veenhuizen et al, 1991; Bobe et al., 2004; Leblanc
et al., 2005). Metabole stoornissen verlagen de productie en verhogen het risico op andere aandoeningen
bv. vruchtbaarheidsstoornissen, zoals dystocie en immuniteitsstoornissen (Veenhuizen et al, 1991; Bobe
et al., 2004). De meeste problemen vinden plaats tijdens de vroege lactatie (Bossaert et al., 2008a;
Bossaert et al., 2008b). Gedurende deze periode is de koe blootgesteld aan verschillende risicofactoren
die zorgen voor stress. Deze factoren kunnen onderverdeeld worden in 3 groepen nl. de verhoging van de
energiebehoefte, de verlaging van de energieopname en de gevoeligheid van het vetweefsel voor
lipolyse. In de praktijk zijn deze factoren steeds sterk met elkaar verweven (Jorritsma et al., 2003;
Bossaert et al., 2008a; Bossaert et al., 2008b).
-de verhoging van de energiebehoefte: Hier hebben we als voorbeeld tweelingdracht en de hoge
melkproductie (Bossaert et al., 2008a).
-de verlaging van de energieopname: Hierbij speelt het management een belangrijke rol. De
voederopname of DSO is zoals, reeds besproken bij 1.2.2 afhankelijk van vele factoren (Nocek en
Braund, 1985; Grummer et al., 2004; Beever, 2006; Bossaert et al., 2008a; Bossaert et al., 2008b).
-de gevoeligheid van het vetweefsel voor lipolyse: Er zijn verschillende factoren die de gevoeligheid van
het vetweefsel voor lipolytische prikkels sterk verhogen en daardoor de bloedconcentratie van NEFA’s
doen toenemen. Eerst en vooral is de lipolytische gevoeligheid gedeeltelijk genetisch vastgelegd, zo zijn
hoogst productieve melkkoeien meer geneigd tot het mobiliseren van vet. Ook de droogstand is belangrijk,
zo is het vet van koeien die in een te ruime conditie afkalven gevoeliger voor lipolyse en minder gevoelig
voor antilypolytische prikkels (Rukkwamsuk et al., 1998; Bossaert et al., 2008a). De overdreven vervetting
bij het afkalven is één van de belangrijkste risicofactoren bij het ontwikkelen van gezondheidsproblemen
(Roche et al., 2009; De Koster en Opsomer, 2012). Daarnaast spelen stress en ziekte ook een rol, deze
zorgen nl. voor een stijging in bloedconcentratie van glucocorticoïden en catecholamines, waardoor er
een verhoogde lipolyse is. De stijging van de concentratie van NEFA’s en/of TG zorgen op hun beurt ook
voor het ontstaan van perifere IR, wat nog versterkt wordt door een hoge concentratie groeihormoon (GH)
aan het begin van de lactatie. De factoren, die dus zorgen voor een verhoging van de lipolyse, werken zo
de functie van insuline tegen (deze remt de o.a. de lipolyse) (Pires et al., 2007; Bossaert et al., 2008a).
Het is niet alleen de aanwezigheid van deze risicofactoren, maar ook de individuele reactie van de koe die
bepaalt of het tot klinische problemen zal komen. Deze individuele adaptatie voor de hoge energievraag
wordt weerspiegeld door de gehalten van NEFA’s voor en na de partus, door accumulatie van TG in de
lever, door het verlies van conditiescore en door persisterende lage bloed glucose en insuline spiegels.
Hiermee wordt in de praktijk dan ook gebruik van gemaakt om de kans op stoornissen in te schatten
Opstapeling van TG in de lever, een persisterende hoge NEFA-concentratie, persisterende lage glucose
en insuline concentraties zijn indicatief voor een zwakke adaptatie (Jorritsma et al, 2001; Jorritsma et al,
2003; Kim en Suh, 2003; Bossaert et al., 2008a).
De grens tussen het fysiologische en pathologische is niet steeds duidelijk daar de ziekten in het
verlengde liggen van de normale adaptatiemechanismen. Metabole stoornissen ontstaan pas wanneer de

15
NEB abnormaal diep is of lang duurt en er een ernstig onevenwicht bestaat tussen de aanvoer en de
verwerkingscapaciteit van NEFA’s van de lever (Bobe et al, 2004; Bossaert et al., 2008a).
Productieziekten hebben vaak een vaag of aspecifiek ziektebeeld, dit komt doordat deze vaak
verschillende orgaanfuncties aantasten. Dit bemoeilijkt dan ook de studies om de exacte oorzaak van een
metabole ziekte te vinden. Vaak liggen dezelfde oorzaken aan de basis van verschillende ziekten bv. bij
ketonemie en leververvetting (zie verder). Ook is het zo dat de meeste gevallen subklinisch verlopen. Het
diagnosticeren en behandelen van metabole stoornissen is in de praktijk dus niet altijd zo simpel. Om toch
een betere houvast te hebben proberen onderzoekers metabole parameters te vinden die rechtstreeks
verbonden zijn met deze aandoeningen. Bijvoorbeeld bij ketonemie: enerzijds hyperketonemie, stijging
van glucagon, NEFA’s, hepatische TG en anderzijds hypoglycemie, hypoinsulinemie en daling van
leverglycogeen en bij leververvetting: verhoogde leverenzymes, ev. leukopenie met linksverschuiving,
verhoogde serumgehaltes van NEFA’s en verlaagde gehaltes van TG en cholesterol en verhoogd TG-
gehalte in de lever. De resultaten van verschillende studies spreken elkaar echter vaak tegen, de
parameters zijn ook vaak zeer variabel waardoor het in de praktijk nog niet echt bruikbaar is (Bobe et al.,
2004; Bossaert et al., 2008a).
2.2. BESPREKING ZIEKTEN
2.2.1 Fat cow syndrome = leververvetting = hepatic lipidosis = fatty liver en ketonemie
Inleiding
Ketonemie en leververvetting gaan vaak hand in hand in de praktijk. Beiden hebben een multifactorieel
karakter, waarbij veranderingen in nutritionele en metabole status een grote rol spelen. Het zijn vnl. de
verstoringen in het lipiden metabolisme, die ontstaan door een verstoring van de homeorhese tijdens de
periode rond de partus, die aan de oorzaak liggen. Er is ook een sterk verband met andere metabole
ziekten, waaronder lebmaag verplaatsing (Wada et al., 1995; Bobe et al., 2004; Hayirli, 2006; Bossaert et
al., 2008a; Bossaert, 2010).
Epidemiologie
Er bestaan verschillende gradaties van leververvetting gaande van een normale lever tot een lever met
milde, matige of erge leververvetting. De onderverdeling en diagnose gebeurt op basis van chemische of
histologische analyse van leverstalen voor lever TG of totaal vet gehalte (Bobe et al., 2004). Hierbij moet
wel een kanttekening gemaakt worden dat koeien verschillend kunnen reageren op eenzelfde TG
concentratie in de lever daar dit indirecte metingen zijn voor het effect op de functies van de levercellen
(Johannsen et al., 1993).
De meeste gevallen van leververvetting vinden plaats in de eerste 4 weken na het kalven (Grummer,
1993; Bobe et al., 2004). In de eerste maand p.p. heeft 30% tot 40% van de koeien te kampen met een
matig vervette lever en 5% tot 10% met een meer ernstige vetinfiltratie, of m.a.w. 50% van de koeien
hebben te maken met leververvetting en daardoor een hoger risico voor andere ziekten en voortplantings-
stoornissen (Bobe et al., 2004). Zoals vermeld kan de lever tot op zekere hoogte een overaanbod aan
NEFA’s verwerken. Door een erg lange of diepe NEB kunnen de koeien in een neerwaartse spiraal
terechtkomen. Ten eerste door de toename van de perifere IR waardoor de lipolyse nog in intensiteit
toeneemt en ten tweede door de aantasting van de leverfunctie (zie figuur 5) (Pires et al., 2007; Bossaert
et al., 2008a).

16
Pathogenese
Leververvetting ontstaat wanneer de opname van vetten door de lever de oxidatie en secretie van vetten
door de lever overschrijdt waardoor de TG dan opgestapeld zullen worden (Rukkwamsuk et al., 1999a;
Herdt, 2000; Bobe et al., 2004; Bossaert et al., 2008a; Bossaert et al., 2008b). Dit is meestal
voorafgegaan door een hoge concentratie aan NEFA’s die zijn vrijgekomen uit het vetweefsel (Bobe et al.,
2004). Histologisch ziet men vetinfiltratie in de cellen, onder de vorm van TG waardoor er zwelling van de
levercellen optreedt en in een vergevorderd stadium necrose met lekkage van enzymen. Macroscopisch
ziet men een graduele zwelling en geelverkleuring van de lever (Bobe et al., 2004; Bossaert et al., 2008a).
Door de leververvetting worden een aantal functies van de lever onderdrukt. Op de eerste plaats wordt de
gluconeogenese verstoord. Hierdoor zullen de glucose- en insulineconcentraties nog verder dalen (zie
figuur 6) (Rukkwamsuk et al., 1999b; Bobe et al., 2004; Murondoti et al., 2004; Bossaert et al., 2008a).
Door deze daling in gluconeogenese is er een gedaalde glucose productie en bijgevolg ook een gedaalde
melkproductie (Bell en Bauman, 1997; De Koster en Opsomer, 2012). De enzymsystemen die instaan
voor de verestering van NEFA’s zijn onderdrukt en er is ook een daling van eiwitten die instaan voor het
verpakken en secreteren van vetten, zoals apoproteine B en proteine kinase C, die belangrijk zijn voor de
redistributie van TG als bouwstenen van de lipoproteinen (Marcos et al., 1990; Katoh, 2002; Bobe et al.,
2004; Bossaert et al., 2008a). Ook wordt een verlaagde activiteit of gevoeligheid van het CPT1
waargenomen en als risicofactor bestempeld. Deze transporteur is verantwoordelijk voor de opname van
NEFA’s in de mitochondriën (zie hoger) (Kräft, 2004; Dann en Drackley, 2005; Bossaert et al., 2008a;
Bossaert et al., 2008b).
Ketonemie en ketoacidose vinden plaats volgend op de leververvetting, nl. doordat de lever de overvloed
aan NEFA’s niet meer kan verwerken en er zo een overvloed aan ketonen ontstaat (Hayirli, 2006;
Bossaert, 2008b).
De aantasting van de leverfunctie gaat ook gepaard met een daling van de eetlust (Bossaert et al.,
2008a).
Risicofactoren
Obesitas (BCS>4.0) bij het afkalven is een van de belangrijkste risicofactoren voor de ontwikkeling van
het fatty liver syndroom. Deze koeien maken nl. een grotere daling in voederopname door en hebben
daardoor een grotere kans op een ergere NEB (Bobe et al., 2004; Drackley et al., 2005; Roche et al.,
2009; De Koster en Opsomer, 2012). Andere factoren die invloed hebben op het ontstaan van
leververvetting zijn o.a. een onvoldoende en onevenwichtig dieet, toxines die het levermetabolisme
beïnvloeden, gestegen oestrogenen gehalte, een gevorderde leeftijd en onvoldoende beweging (Bobe et
al., 2004).
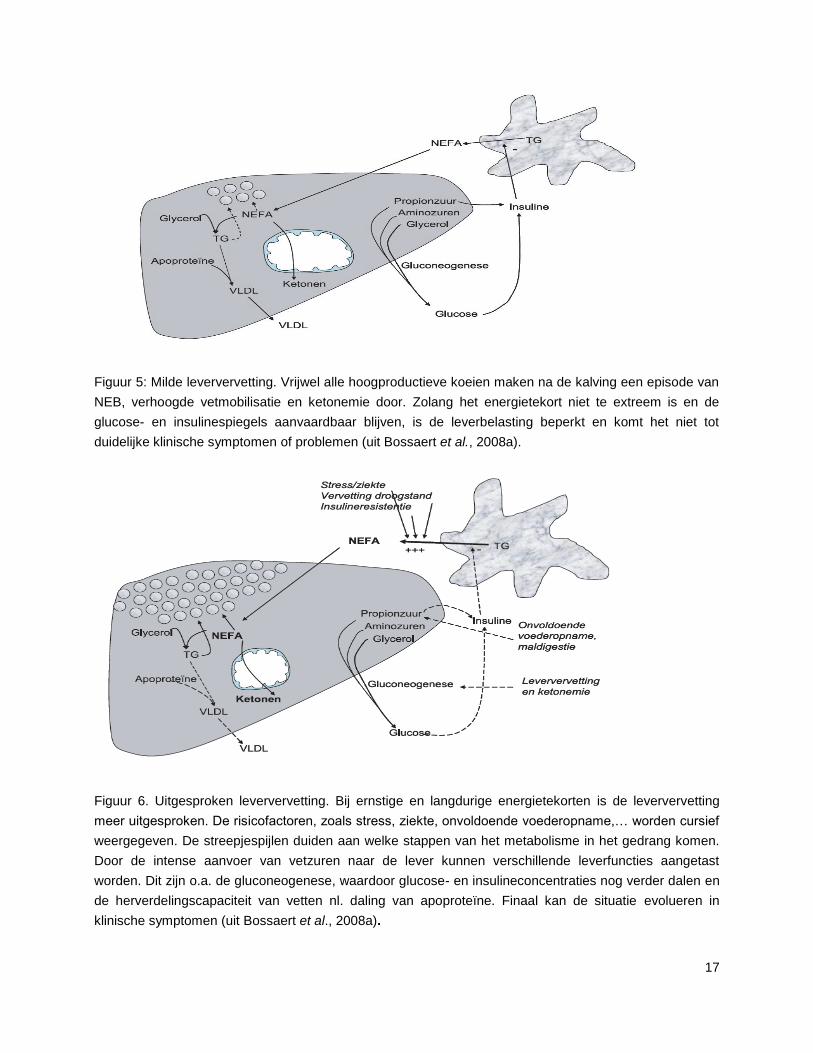
17
Figuur 5: Milde leververvetting. Vrijwel alle hoogproductieve koeien maken na de kalving een episode van
NEB, verhoogde vetmobilisatie en ketonemie door. Zolang het energietekort niet te extreem is en de
glucose- en insulinespiegels aanvaardbaar blijven, is de leverbelasting beperkt en komt het niet tot
duidelijke klinische symptomen of problemen (uit Bossaert et al., 2008a).
Figuur 6. Uitgesproken leververvetting. Bij ernstige en langdurige energietekorten is de leververvetting
meer uitgesproken. De risicofactoren, zoals stress, ziekte, onvoldoende voederopname,… worden cursief
weergegeven. De streepjespijlen duiden aan welke stappen van het metabolisme in het gedrang komen.
Door de intense aanvoer van vetzuren naar de lever kunnen verschillende leverfuncties aangetast
worden. Dit zijn o.a. de gluconeogenese, waardoor glucose- en insulineconcentraties nog verder dalen en
de herverdelingscapaciteit van vetten nl. daling van apoproteïne. Finaal kan de situatie evolueren in
klinische symptomen (uit Bossaert et al., 2008a).

18
Kliniek
Zoals vermeld zijn de symptomen van metabole ziekten vaak vaag en zijn er veel subklinische gevallen,
dit is hier ook het geval voor beide ziekten. Zelden zijn er duidelijke symptomen zoals hypofagie, een
sterke daling van de melkproductie, een groot gewichtsverlies, lethargie en hyperexcitatie. Soms kan de
situatie verergeren en evolueren naar nerveuze ketonemie of hepato-encephalopathie, hierbij zien we dan
vnl. (centrale) zenuwstoornissen zoals gedaald bewustzijnsniveau, ataxie en coma en eventuele sterfte
door lever- en nierfalen of hartstilstand (Rehage et al., 1999; Bobe et al., 2004; Bossaert et al., 2008a).
Onder vage symptomen verstaan we het volgende:
1) Er is een daling van de melkproductie. Dit komt deels door een gedaalde eetlust, maar in belangrijke
mate door de onderdrukking van de gluconeogenetische capaciteit van de lever. Door de gedaalde
glucose productie zal er minder melk geproduceerd worden (Baird, 1982; Bobe et al., 2004; Hayirli,
2006; Bossaert et al., 2008a; De Koster en Opsomer, 2012).
2) Ketonemie en leververvetting hebben steeds de neiging om te verergeren en te evolueren naar andere
ziekten. Dit is te wijten aan de gedaalde eetlust, de chronische lage insuline spiegels, de perifere IR en
de mogelijk verminderde activiteit van enzymen en cofactoren die instaan voor de gluconeogenese, de
mitochondriale opname van vetzuren en de redistributie van TG (Van Winden et al., 2003a en b;
Hayirli, 2006; Bossaert et al., 2008a).
3) Er is ook een algemene immuniteitsdaling. Dit wordt veroorzaakt door de hoge gehalten aan
ketolichamen en NEFA’s, die zowel het aantal als de chemotaxis, de fagocytose en de
cytokineproductie van de leukocyten kunnen onderdrukken. Hierdoor zijn de dieren vatbaarder voor
infecties en ontstekingen zoals bv. mastitis en metritis (Suriyasathaporn, 2000; Ohtsuka et al., 2001,
Bobe et al., 2004; Hayirli, 2006; Scalia et al., 2006).
4) De koeien hebben ook een gedaald voortplantingsvermogen, dit o.a. door een aantasting van de
follikel, eicel, embryo en baarmoeder. Hierdoor is er o.a. een verhoogde kans op dystocie. Dit zou
veroorzaakt worden door een gedaalde en vertraagde synthese van steroïdhormonen nl. progesteron
en luteïniserend hormoon (LH), een erge NEB, een gedaalde concentratie van insulin-like growth
factor 1 (IGF-I), insuline en lipoproteïnen en verhoogde gehaltes aan NEFA’s, ammoniak en ureum
(Veenhuizen et al.,1991; Bobe et al., 2004; Leroy et al., 2005; Vanholder et al., 2005; Hayirli, 2006).
Deze vage en subklinische gevallen gaan gepaard met economische schade en dienen dus zo goed
mogelijk behandeld te worden, zelfs nog beter vermeden te worden door preventief te handelen. Deze
schade is o.a. te wijten aan de verhoogde diergeneeskundige kosten, een langere tussenkalftijd, een
verminderde melkproductie en een verkorte levensverwachting van de koeien (Bobe et al., 2004; Bossaert
et al., 2008a).
Preventie en therapie
De nadruk zou dus moeten liggen op preventie i.p.v. behandeling. Deze preventie wordt bekomen door
het voeren van een goede bedrijfsmanagement, dit bestaat o.a. uit het voorzien van voldoende goede
voeding en zorgen voor een propere en stressloze omgeving in de periode rond het afkalven. Deze
maatregelen zijn in sommige gevallen niet voldoende. Bij de preventie en behandeling ligt de nadruk op
het verhinderen van een overdreven lipolyse van het vetweefsel, een verbetering van de afvoer van TG
vanuit de lever, het voldoende doen toenemen van de gluconeogenese van de lever of de glucose
voorzieningen en de glucose opname van de extra-hepatische weefsels doen toenemen (Bobe et al.,
2004; Hayirli, 2006). Een mogelijke preventieve actie is het drenchen met propyleenglycol in de periode
rond de partus, dit zorgt nl. voor een toegenomen plasmaglucose en insuline concentratie en een

19
gedaalde plasma BHB en NEFA concentratie (Studer et al., 1993; Christensen et al., 1997; Duffield, 2000;
Pickett et al., 2003). Een andere mogelijkheid is het injecteren van glucocorticoïden, glucagon of lage
dosissen van slow-release insuline in de eerste dagen na het kalven, deze zijn echter enkel theoretisch
daar glucagon en insuline niet mogen gebruikt worden bij melkvee en de toediening van glucocorticoïden
ook een groot risico met zich meebrengt (Hayirli et al., 2002; Nafikov et al., 2002; Bobe et al., 2004; Cools
et al., 2008b). Hierbij dient toch opgemerkt worden dat er variatie is tussen de resultaten van verschillende
proeven, zo is het effect afhankelijk is van de dosis en de toedieningsvorm en is er nog verder onderzoek
vereist (Bobe et al., 2004; Cools et al., 2008b).
De behandeling van leververvetting en/of ketonemie bestaat in de eerste plaats uit het reduceren van de
NEB en de koe voldoende voorzien van glucose voor de melkproductie (Cools et al., 2008b). Hierbij is het
voorzien van een goede energierijke en uitgebalanceerde voeding en omgeving het belangrijkste(Bobe et
al., 2004; Cools, 2008b). Daarnaast zijn nog andere zaken mogelijk als extra ondersteuning zoals o.a.
oraal drenchen van glucose precursoren of een intraveneus glucose infuus eventueel ondersteund met
een éénmalige injectie corticoïden, maar vele van deze technieken moeten nog verder uitgetest worden
(Bobe et al., 2004; Cools, 2008b).
2.2.2 Lebmaagverplaatsing
Een lebmaagverplaatsing is een veel voorkomende aandoening bij hoogproductief melkvee (Türck en
Leonhard-Marek, 2010). De lebmaag kan naar links (LDA= left displacement abomasum) of naar rechts
verplaatsen. Verder zijn 2 types lebmaagverplaatsing te onderscheiden: een klassieke en atypische
verplaatsing. De klassieke lebmaag verplaatsing komt vnl. bij het hoogproductief melkvee voor en heeft
meestal een functionele onderliggende oorzaak. De exacte oorzaak is nog niet volledig opgehelderd maar
mogelijke oorzaken zouden zijn: een te hoog NEFA gehalte, een te zure inhoud in het duodenum,
hypocalcemie, ontstekingsprocessen, hypokaliëmie en hyperinsulinemie (Doll et al, 2009; Türck en
Leonhard-Marek, 2010; Deprez en van Loon, 2012). Een toegenomen extracellulaire concentratie van
insuline heeft invloed op de contractie activiteit van de spieren van de lebmaag. De effecten van een te
hoog insuline gehalte kunnen ongedaan worden door barium, glibenclamide of ouabain. Het
onderliggende mechanisme zou een toegenomen K+
geleiding of een toegenomen activiteit van Na/K-
ATPase of beide zijn (Türck en Leonhard-Marek, 2010).
Men ziet ook dat lebmaagverplaatsingen veel voorkomen bij koeien met een vervette lever of ketose, zelfs
in het subklinisch stadium (Van Winden et al., 2003a en b). Hierbij werd in de periode kort voor de
klinische diagnose van de verplaatsing verlaagde bloedconcentraties van calcium, glucose en insuline en
verhoogde concentraties van NEFA’s, BHB en aspartaat aminotransferase activiteit vastgesteld. Deze
parameters spelen een belangrijke rol in de pathogenese van LDA (Van Winden et al., 2003a). Dus zowel
een verhoogde concentratie als een verlaagde concentratie van insuline kan een rol spelen in de
pathogenese van lebmaagverplaatsing (Van Winden et al., 2003a; Türck en Leonhard-Marek, 2010). Om
het risico op het krijgen van LDA in te schatten zien we dat een bepaling van NEFA en BHB in de laatste
week prepartum indicatief blijken te zijn (Leblanc et al., 2005).
De atypische vorm komt voor bij stieren, vaarzen en vleeskoeien, is meestal accidenteel en heeft meestal
een mechanische oorzaak zoals een hypertrofie of afsnoering van de pylorus of afsnoering van de dunne
darm, ulcer, vergroeiingen,… (Deprez en van Loon, 2012).

20
2.2.3 Andere metabole ziekten
Naast leververvetting, ketonemie en lebmaagverplaatsing zijn er ook nog een aantal andere metabole
ziekten waarmee hoogproductieve dieren vaak te kampen hebben. Deze zijn o.a. endometritis, metritis,
pyometra, cysteuze ovariële follikels, ketonemie, mankheid, mastitis, milk fat depression, hypocalcemie,
pensverzuring of Sub-Acute Ruminal Acidosis (SARA), retentio secundinarum of aan de nageboorte
blijven staan en uier oedeem (Heuer et al. Bossaert et al., 2008a). Tussen deze ziekten zijn er vaak
onderlinge linken. In de studie van Heuer et al. (1999) bijvoorbeeld werd gekeken naar de relatie tussen
de BCS en de melk gegevens van de 1e test dagen en de incidentie van ziektes, melkproductie en
opruiming. Gedurende deze studie werd een wegen-model analyse met aanpassing voor tijd en risico
toegepast om de opeenvolging van de gebeurtenissen te benadrukken, dit is te zien op figuur 7.
Figuur 7. Wegen-model analyse van relatie tussen de BCS en de melk gegevens van de 1e test dagen en
de incidentie van ziektes, melkproductie en opruiming.
De pijlen die variabelen verbinden met een box duiden een verband aan met alle variabelen binnen de
box. Tweerichtingspijlen duiden een individuele relatie aan in beide richtingen.
Variabelen met hun afkortingen: Endo ≤20 d = endometritis, metritis, of pyometra 1e 20 dagen post partum
(p.p).; Endo >20 d = endometritis, metritis, of pyometra langer dan 20dagen p.p.; Cyst = cystic ovariële
ziekte; Ket = ketosis of ketonemie; Lame = lameness = mankheid; LDA = left (mostly) displaced
abomasum = linker (meest) lebmaagverplaatsing; Mast = mastitis; MF = milk fever = hypocalcemie; RP =
retained placenta= retentio secundinarum= aan de nageboorte blijven staan; Preg1 = eerste
bevruchtingsrate; FP1 >1.5 = vet-eiwit (fat to protein) ratio van de 1e test dagen p.p.>1.5 ; M1 =
melkproductie van de 1e dag p.p.; BCS1 ≥4 = eerste BCS p.p. ≥4; BCS1 ≤2 = eerste BCS p.p. ≤2; BCS
loss = verschil tussen de eerste en tweede BCS p.p.>0.5; CC = interval tussen kalving tot bevruchting
(calving to conception); SPC = services per conception; FCM100 = 100 dagen melkgift, gecorrigeerd voor
4%vet en 3.2% eixit ; and Cull = culling rate= opruimings getal.
De asteriks duidt het verband aan tussen de retentio secundinarum (RF) en hypocalcemie (MF) (uit Heuer
et al., 1999).
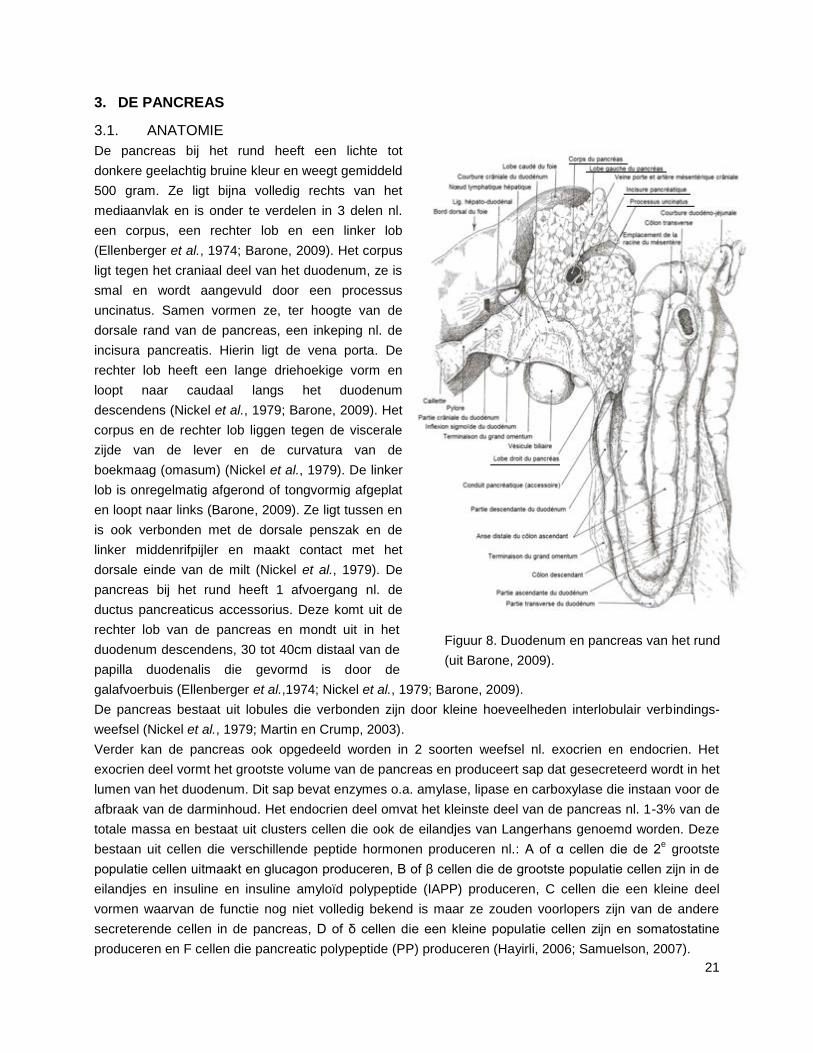
21
Figuur 8. Duodenum en pancreas van het rund
(uit Barone, 2009).
3. DE PANCREAS
3.1. ANATOMIE
De pancreas bij het rund heeft een lichte tot
donkere geelachtig bruine kleur en weegt gemiddeld
500 gram. Ze ligt bijna volledig rechts van het
mediaanvlak en is onder te verdelen in 3 delen nl.
een corpus, een rechter lob en een linker lob
(Ellenberger et al., 1974; Barone, 2009). Het corpus
ligt tegen het craniaal deel van het duodenum, ze is
smal en wordt aangevuld door een processus
uncinatus. Samen vormen ze, ter hoogte van de
dorsale rand van de pancreas, een inkeping nl. de
incisura pancreatis. Hierin ligt de vena porta. De
rechter lob heeft een lange driehoekige vorm en
loopt naar caudaal langs het duodenum
descendens (Nickel et al., 1979; Barone, 2009). Het
corpus en de rechter lob liggen tegen de viscerale
zijde van de lever en de curvatura van de
boekmaag (omasum) (Nickel et al., 1979). De linker
lob is onregelmatig afgerond of tongvormig afgeplat
en loopt naar links (Barone, 2009). Ze ligt tussen en
is ook verbonden met de dorsale penszak en de
linker middenrifpijler en maakt contact met het
dorsale einde van de milt (Nickel et al., 1979). De
pancreas bij het rund heeft 1 afvoergang nl. de
ductus pancreaticus accessorius. Deze komt uit de
rechter lob van de pancreas en mondt uit in het
duodenum descendens, 30 tot 40cm distaal van de
papilla duodenalis die gevormd is door de
galafvoerbuis (Ellenberger et al.,1974; Nickel et al., 1979; Barone, 2009).
De pancreas bestaat uit lobules die verbonden zijn door kleine hoeveelheden interlobulair verbindings-
weefsel (Nickel et al., 1979; Martin en Crump, 2003).
Verder kan de pancreas ook opgedeeld worden in 2 soorten weefsel nl. exocrien en endocrien. Het
exocrien deel vormt het grootste volume van de pancreas en produceert sap dat gesecreteerd wordt in het
lumen van het duodenum. Dit sap bevat enzymes o.a. amylase, lipase en carboxylase die instaan voor de
afbraak van de darminhoud. Het endocrien deel omvat het kleinste deel van de pancreas nl. 1-3% van de
totale massa en bestaat uit clusters cellen die ook de eilandjes van Langerhans genoemd worden. Deze
bestaan uit cellen die verschillende peptide hormonen produceren nl.: A of α cellen die de 2e grootste
populatie cellen uitmaakt en glucagon produceren, B of β cellen die de grootste populatie cellen zijn in de
eilandjes en insuline en insuline amyloïd polypeptide (IAPP) produceren, C cellen die een kleine deel
vormen waarvan de functie nog niet volledig bekend is maar ze zouden voorlopers zijn van de andere
secreterende cellen in de pancreas, D of δ cellen die een kleine populatie cellen zijn en somatostatine
produceren en F cellen die pancreatic polypeptide (PP) produceren (Hayirli, 2006; Samuelson, 2007).

22
3.2. ROL VAN DE ENDOCRIENE PANCREAS
3.2.1. Inleiding
De controle van het metabolisme tijdens de dracht en lactatie is gebaseerd op twee belangrijke principes:
homeostase en homeorhese.
Homeostase wordt beschreven als “acute endocriene reacties die plaatsvinden binnen het lichaam om
bepaalde fysiologische en metabole parameters, zoals bijvoorbeeld de lichaamstemperatuur en de
glucosespiegel, binnen nauwe grenzen te houden”.
Daartegenover staat homeorhese, beschreven als: “het samenspel van metabole veranderingen in het
lichaam om een gewijzigde fysiologische status, zoals de overgang van droogstand naar lactatie, te
ondersteunen. Hieronder zit onder andere de regeling van verdeling van voedingstoffen gedurende de
dracht en de lactatie bv. lipolyse en lipogenese”. Homeorhese is, in tegenstelling tot homeostase, eerder
chronisch, omvat meerdere hormonen, beïnvloedt meerdere orgaansystemen en veroorzaakt vaak ook
een gewijzigde biologische respons op endocriene signalen. De aanpassingen tijdens de lactatiecyclus
zoals de koe tijdens de transitieperiode zich moet aanpassen aan de overschakeling vrucht naar
melkproductie, de lactatiepiek en de droogstand voldoet aan al deze criteria en is dus een
schoolvoorbeeld van homeorhese (Bauman en Currie, 1980; Bell, 1995; Bossaert et al., 2008b).
Hierna wordt de rol, in de homeorhese, van de verschillende hormonen geproduceerd door de endocriene
pancreas besproken. Deze hormonen zijn insuline, glucagon, somatostatine, PP en IAPP (Hayirli, 2006;
Samuelson, 2007).
3.2.2. Insuline
Algemeen
Insuline is een anabool hormoon, het heeft een stapelfunctie en speelt een belangrijke rol in het
metabolisme van koolhydraten, vetten en eiwitten. Hierbij speelt glucose een centrale rol en heeft insuline
een grote invloed op de werking van de lever, de skeletspieren en het vetweefsel (Brockman en Laarveld,
1986; Opsomer, 1995; Frandson, 2003; van der Putten, 2005; Hayirli, 2006). Insuline verlaagt de bloed
glucose spiegel, voornamelijk door de cellulaire opname van glucose in skeletspieren en vetweefsel te
stimuleren en door inhibitie van glucose aanmaak (gluconeogenese) en van glucose vrijstelling uit
glycogeen, de stockage vorm van glucose (glycogenolyse) door de lever (Brockman en Laarveld, 1986;
Herdt, 1988; Torck, 1993; Samuelson Don, 2007). Naast deze invloeden worden in de lever de
ketogenese en de eiwit afbraak geremd en de afbraak van glucose (glycolyse), de opbouw van glycogeen
(glycogenese), de eiwit synthese en de TG synthese gestimuleerd (Brockman en Laarveld, 1986;
Frandson et. al., 2003; Hayirli, 2006). In het vetweefsel zorgt het hormoon ook voor een stimulatie van de
vetaanmaak (lipogenese) en een inhibitie van de vetafbraak (lipolyse) (Brockman en Laarveld, 1986;
Opsomer, 1995; Frandson et. al., 2003). Ter hoogte van het spierweefsel remt insuline ook de eiwit
afbraak (proteolyse) en de glycogenolyse en stimuleert het de proteïnesynthese, het gebruik van ketonen,
de glycolyse en de glycogenese (Brockman en Laarveld, 1986; Opsomer, 1995; Frandson et. al., 2003;
Hayirli, 2006).
Naast de verschillende invloeden op celniveau, heeft insuline ook een invloed op de eetlust, nl. een
verlaagd insuline gehalte zorgt voor een toegenomen eetlust (dus insuline remt de eetlust). Dit effect
wordt op lange termijn ook deels gezien door GH stijgingen en wordt gezien tijdens de vroege lactatie wat
zorgt voor een geleidelijke toename van DSO na de kalving (McDowell, 1991; Bossaert et al., 2008b).

23
Om actief te zijn moet insuline binden op een receptor, een glycoproteïne. De binding zal zorgen voor
signaaltransductie binnen de cel, dit is zeer complex en kan omschreven worden in 3 fasen nl.:
(auto)fosforylatie van de insuline receptor, gevolgd door intracellulaire signaal overbrenging door de
second messenger en tenslotte de translocatie van de glucose transporters (GLUT) (Saul, 2000; Hayirli,
2006). De opname van glucose door de cellen gebeurt dus finaal door gefaciliteerd transport via
membraan-gebonden GLUT. Er zijn verschillende GLUT-typen die allen op verschillende manieren tot
expressie gebracht worden op het celoppervlak. Deze verschillende posities zorgen ervoor dat er een
verschillende weefsel respons is van insuline voor de opname van glucose. Zo is geweten dat GLUT 1
vnl. voorkomt in de hersenen, de placenta, de uier en de rode bloedcellen; GLUT 2 in de lever, de nier en
de pancreas; GLUT 3 in de hersenen en placenta; GLUT 4 in het vetweefsel, skeletspieren en hartspier;
en GLUT 5 in de dunne darm. Enkel GLUT 4 is insuline afhankelijk en heeft insuline nodig voor de
opname van glucose. De andere receptoren, zoals GLUT 1 in de uier en de placenta zijn insuline
onafhankelijk en hebben geen insuline nodig voor de opname van glucose. Dit mechanisme speelt een
belangrijke rol in het glucose metabolisme (Zhao et al., 1996; Hayirli, 2006; Zhao and Keating, 2007;
Bossaert et al., 2008b).
Er zijn veel zaken die de insuline vrijstelling beïnvloeden. Zaken die de vrijstelling stimuleren zijn
voedingsstoffen of m.a.w. metabolieten van het koolhydraat-, vet- en eiwitmetabolisme (waaronder een
verhoogd glucose gehalte als belangrijkste factor, galactose, glyceraldehyde, lysine, leucine, alanine,
lange-keten vetzuren, kalium en calcium), gastro-intestinale hormonen (o.a. glucagon, PP, ‘gastric
inhibitory polypeptide’, secretine en cholecystokine), parasympatische stimuli (o.a. vagus activiteit, beta-
adrenerge activiteit en acetylcholine), medicatie (o.a. sulfonamide) en een reeks andere hormonen (GH,
adrenocorticotroop hormoon (ACTH), Thyroïd stimulerend hormoon (TSH) en enkele steroïdhormonen).
Zaken die de vrijstelling inhiberen zijn fysiologische condities (o.a. vasten en training), gastrointestinale
hormonen (o.a. somatostatine, pancreastatin, galanin), sympatische stimuli (adrenerge activiteit bv. stress
met vrijstelling van o.a. catecholamines) en andere specifieke stoffen (o.a. interleukine-1 (IL-1) en
prostaglandine F2-alfa (PGF2-alfa)) en NEFA’s (Hayirli, 2006; Pires et al., 2007; Silbernagl en
Despopoulos, 2008; Berne et al., 2010).
Men ziet ook dat het insulin metabolisme een genetische achtergrond heeft. Zo ziet men dat koeien die
geselecteerd zijn op een hogere melkproductie een grotere IR hebben en verondersteld men dat
hoogproductieve koeien de predispositie hebben op verlaagde insuline spiegels (Swali en Wathes, 2006;
Chages et al., 2009).
Tijdens de periode van late dracht en tijdens de partus daalt het gehalte aan insuline in het bloed en is er
verminderde gevoeligheid van de perifere weefsels, dit veroorzaakt de belangrijkste homeorhetische
aanpassingen (Bossaert et al., 2008b; Hermans, 2010; Schoenberg en Overton, 2010). De daling van het
insuline gehalte is het resultaat van de lage glucosespiegel en van een tijdelijke onderdrukking van de
endocriene pancreasfunctie wat resulteert in een verminderde secretie (Holtenius et al., 2003; Bossaert et
al, 2008b; Bossaert et al., 2008c). Naast de daling van insuline zijn er ook nog andere hormonale
veranderingen die de effecten van het laag insuline gehalte versterken, zo is er een stijging in GH en een
daling van IGF-1, een verhoging in catecholamines en een verhoging in glucocorticoïden (Opsomer 1995;
Zulu et al., 2002; Bossaert et al., 2008b). Deze veranderingen zorgen er globaal voor dat er een
verhoogde lipolyse is t.h.v. het vetweefsel, er een daling is van de proteïnesynthese t.h.v. het spierweefsel
en de gluconeogenese en de ketogenese die plaatsvinden in de lever gestimuleerd worden, waardoor

24
bloedglucosespiegel op peil wordt gehouden en de ketonen als energiebron gebruikt kunnen worden door
perifere weefsels (Opsomer, 1995; Samuelson Don, 2007; De Koster en Opsomer, 2012).
Rol in de productie
Insuline speelt een belangrijke rol in de energiebalans en de melkproductie bij het hoogproductief melkvee
(Opsomer, 1995; Bossaert et al., 2008b; Bossaert, 2010).
Door de specifieke verdeling van de GLUT kan de melkklier tijdens insulinearme omstandigheden
onbeperkt glucose opnemen, soms tot 85% van alle bloedglucose, terwijl het perifere glucose verbruik tot
een minimum beperkt wordt (Knight et al., 1994; Zhao et al., 1996). Dit is belangrijk daar, zoals reeds
eerder vermeld, de groeiende vrucht en de uier bijna uitsluitend aangewezen zijn op glucose. Tijdens de
toplactatie wordt daarom al het glucose voorbehouden voor de uier en de opname t.h.v. de perifere
weefsels zoveel mogelijk beperkt, zie figuur 9. Dit in tegenstelling tot het verdere verloop van de lactatie
en de droogstand waarbij, samen met de gestegen voederopname, de insulineconcentratie zal toenemen
en de perifere weefsels wel gebruik kunnen maken van glucose, zie figuur 10 (Opsomer, 1995; Bossaert
et al., 2008b; Bossaert, 2010; Schoenberg en Overton, 2010).
Figuur 9. De glucosehuishouding tijdens de lactatie (negatieve energiebalans).
Tijdens de lactatie is er een laag gehalte aan insuline en zijn de expressie van de insuline-onafhankelijke
GLUT 1-2 (glucosetransporter molecule) van de uier verhoogd en die van de insuline afhankelijke GLUT 4
in de perifere weefsels zoals vet en spieren verlaagd wat ervoor zorgt dat uier onbeperkt glucose kan
opnemen en de perifere weefsels aangewezen zijn op andere bronnen zoals ketonen en vetzuren als
energiebron. In de lever wordt uit propionzuur uit het dieet en aminozuren afkomstig van de spieren
glucose gesynthetiseerd. De lage gehaltes insuline zorgen voor een hogere lipolyse. De NEFA’s die
vrijkomen worden geoxideerd in de mitochondria en zullen tijdens de NEB voornamelijk aanleiding geven
tot de vorming van ketonen, die zoals vermeld kunnen gebruikt worden als energiebron in de perifere
weefsels (uit Bossaert et al., 2008b; Bossaert, 2010).

25
Figuur 10. De glucosehuishouding tijdens de droogstand (positieve energiebalans).
Bij voldoende insuline komen de insulineafhankelijke glucosetransporters (GLUT4) tot expressie op de
spieren en het vetweefsel waardoor ze, naast de lever, glucose kunnen opnemen. Door de hoge gehaltes
aan insuline vindt de lipolyse plaats aan een lage snelheid; de circulerende NEFA’s worden opgenomen
door de lever en worden terug opgeslagen in het vetweefsel onder de vorm van VLDL, meer dan te
worden geoxideerd in de mitochondria (uit Bossaert et al., 2008b; Bossaert, 2010).
Naast insuline zorgt de gestegen GH concentratie voor een verhoogde bloedtoevoer naar de uier wat de
melkproductie ook stimuleert (Knight et al., 1994; Zhao et al., 1996; Bossaert et al, 2008b).
Zoals reeds eerder vermeld is de aanvoer van propionzuur, de belangrijkste glucogene bouwsteen voor
de gluconeogenese in de lever bij herkauwers, vanuit de pens gelimiteerd en is er nood aan andere
glucogene bouwstenen nl. spiereiwitten en in beperkte mate glycerol. Hierbij speelt insuline een
belangrijke rol, een daling in insuline zorgt nl. voor een stijging van de aanvoer van glucogene
bouwstenen en de gluconeogenese in de lever. De gestegen GH concentraties stimuleren dit ook
(Bossaert et al., 2008b; Bossaert, 2010).
De lipolyse, die zoals reeds eerder vermeld tijdens de NEB plaats vindt, wordt ook sterk beïnvloed door
insuline. Het enzym dat verantwoordelijk is voor de lipolyse, nl. het hormoonsensitive lipase, wordt
geïnhibeerd door insuline en gestimuleerd door glucagon, catecholaminen en glucocorticoïden. Bij lage
insulinespiegels tijdens de NEB heeft de lipolyse dus de overhand wat finaal zorgt voor het ontstaan van
ketonen, die kunnen gebruikt worden in de perifere weefsels als brandstof (McNamara en Hillers, 1986;
Kokkonen, 2005; Bossaert, 2010).
Daar insuline ook een invloed heeft op de DSO is ze dus ook zeer belangrijk, daar dit meestal een
limiterende factor is in de productie (Bossaert et al., 2008b).
Finaal kan men dus besluiten dat het lage gehalte aan insuline een grote rol heeft in het energie-
metabolisme en de glucose aanvoer naar de uier tijdens de toplactatie wat ervoor zorgt dat er zeer grote
hoeveelheden melk kunnen geproduceerd worden (Bossaert et al., 2008b).

26
Rol in de gezondheid
Insuline speelt ook een belangrijke rol in de pathogenese van verschillende metabole ziekten, hier zal de
rol in de meest voorkomende ziekten besproken worden.
De rol van insuline bij de leververvetting en ketonemie speelt zich af op verschillende vlakken, nl. in de
pathogenese en de preventie en behandeling (Bobe et al., 2004; Hayirli, 2006; Bossaert et al., 2008a en
b; Cools et al., 2008b).
Zoals reeds vermeld, vindt er tijdens de NEB lipolyse door een gedaalde insuline gevoeligheid en gedaald
insuline gehalte plaats waarbij NEFA’s vrijkomen, die vervolgens verwerkt worden in de lever en daar
kunnen leiden tot een mildere tot erge leververvetting (Rukkwamsuk et al., 2000; Bossaert et al., 2008b).
Leververvetting is geassocieerd met een gewijzigde hormoon gevoeligheid van het vetweefsel, zo is bv.
de lipogenese minder gestimuleerd door insuline en glucose (Rukkwamsuk et al., 1999c) en is de lipolyse
minder geremd door glucose en BHB (Rukkwamsuk et al., 1998). Daarnaast is er ook associatie met de
veranderingen in de concentratie van de hormonen zelf. Dit komt door de gedaalde klaring van hormonen,
zoals insuline uit levercellen die geïnfiltreerd zijn met vetten. Dit zorgt waarschijnlijk voor een daling in de
productie of secretie t.h.v. de pancreas (Strang et al., 1998; Bobe et al., 2004).
Door de leververvetting worden een aantal functies van de lever dus onderdrukt, waardoor de glucose- en
insulineconcentraties nog verder dalen (Rukkwamsuk et al.,1999b; Bobe et al., 2004;Bossaert et al.,
2008a). Dit is negatief daar dit zal samen gaan met een gedaalde melk productie (Bell en Bauman, 1997;
De Koster en Opsomer, 2012), het verergeren van de leververvetting en het ontstaan van andere ziekten
(Bobe et al., 2004; Bossaert, 2008a)
Het insuline metabolisme speelt ook een belangrijke rol in de preventie en behandeling van de
leververvetting en ketonemie. Hierbij ligt nl. de nadruk op het verhinderen van een overdreven lipolyse,
een verbetering van de afvoer van TG vanuit de lever, het voldoende doen toenemen van de
gluconeogenese van de lever of de glucose voorziening, en de glucose opname van de extra-hepatische
weefsels doen toenemen (Bobe et al., 2004; Hayirli, 2006). Dit kan indirect via het drenchen met
propyleenglycol in de periode rond de partus, dit zorgt o.a. voor een toegenomen plasmaglucose en
insuline concentratie (Studer et al., 1993; Christensen et al., 1997; Duffield, 2000; Pickett et al., 2003).
Een andere mogelijkheid is het injecteren van lage dosissen van slow-release insuline in de 1e dagen na
het kalven, dit is echter enkel theoretisch daar insuline niet mag gebruikt worden bij melkvee (Hayirli et al.,
2002; Bobe et al., 2004; Cools et al., 2008b).
De rol in de pathogenese van lebmaagverplaatsing is dubbel, zo ziet men dat zowel een verhoogde als
verlaagde concentratie de ziekte kunnen veroorzaken (zie 2.2.2) (Van Winden et al., 2003a; Türck en
Leonhard-Marek, 2010).
Insuline speelt ook een rol bij de ontwikkeling van cysteuze ovariële follikels (Opsomer et al., 1999). Er
is reeds aangetoond dat er insuline receptoren aanwezig zijn de ovaria van runderen. Insuline stimuleert
de estradiol en androsteendion productie door de follikels (Spicer et al., 1993; Spicer en Echternkamp,
1995; Butler et al., 2004). Ook stimuleert insuline de rijping van de follikels en speelt ze een rol in de
selectie van de dominante follikel en heeft dus een belangrijke functie in de oestrus cyclus (Kawashima et
al., 2007; Butler et al., 2004). Daar insuline belangrijk is in de folliculaire ontwikkeling is een verlaagd
gehalte geassocieerd met de vorming van cysten (Vanholder et al., 2005; Braw-Tal et al., 2009).

27
Insuline resistentie en secretie
Zoals hoger vermeld worden de effecten van insuline geregeld door enerzijds de insuline secretie door de
pancreas en anderzijds door de insuline gevoeligheid van de weefsels.
Insuline resistentie (IR)
Gedurende de laatste 2 decennia heeft het concept van IR veel aandacht gekregen in het onderzoek bij
de mens door de toegenomen prevalentie van obesitas en de geassocieerde ziekten zoals type 2
diabetes en cardiovasculaire aandoeningen. Deze worden samengenomen onder de term ‘metabool
syndroom’ (Cornier et al., 2008; Schoenberg en Overton, 2010).
Insuline resistentie beschrijft de toestand in welke een fysiologisch gehalte van insuline een lagere dan
normale biologische respons vertoont. Deze omvat veranderingen in de gevoeligheid (de hoeveelheid van
een hormoon die nodig is om een half maximaal effect uit te lokken) of de respons (het maximale effect
van insuline) of beide (Kahn, 1978; Sano et al., 1991). Het moleculair mechanisme van IR kan
gelokaliseerd zijn op 3 niveaus nl.: pre-receptor level, wat een gedaalde insuline productie, toegenomen
insuline degradatie of beide omvat; receptor level, wat een gedaald nummer van receptoren en een
gedaalde bindingsaffiniteit omvat; en post-receptor level, wat een ongelijke intracellulaire respons en een
translocatie van GLUT omvat. Algemeen ziet men dat defecten t.h.v. het pre-receptor level
hypoinsulinemie veroorzaken, t.h.v. het receptor level verminderde insuline repons veroorzaken en
defecten t.h.v. het post-receptor level verminderde insuline gevoeligheid veroorzaken (Kahn, 1978).
Er zijn verschillende zaken die een invloed hebben op het ontstaan van IR. Deze factoren zijn o.a.
ontwikkelende dracht, obesitas, hyperinsulinemie, voeding van vet, malnutrition en andere hormonen
(Hayirli, 2006).
Daar de uterus en de uier bij de herkauwers bijna uitsluitend zijn aangewezen op glucose als energiebron
worden tijdens het einde van de dracht en nog meer tijdens de vroege lactatie de perifere weefsels, nl. het
vet- en spierweefsel meer IR en is er een laag plasma insuline gehalte. Dit is mogelijk dankzij de
verschillen in GLUT-typen. Er treedt een ‘glucose sparend’ effect op waarbij er de voorkeur van nutrienten
naar de dracht en melkproductie gaat. Dit is fysiologisch tijdens de vroege lactatie (Bell, 1995; Bell en
Bauman, 1997; Opsomer, 1995; Hayirli, 2006; Bossaert et al., 2008a en b; Schoenberg en Overton,
2010).
Door de perifere IR is er in de vroege lactatie altijd een toegenomen lipolyse en een toegenomen NEFA
plasmaconcentratie aanwezig (Pires et al., 2007; Opsomer, 1995; Bossaert et al., 2008a; Schoenberg en
Overton, 2010). Door een erg lange of diepe NEB kunnen de koeien in een neerwaartse spiraal
terechtkomen. Dit resulteert nl. in een zeer grote vetmobilisatie wat zorgt voor een grote overvloed aan
NEFA’s. Deze veroorzaken, zoals eerder vermeld, o.a. een toename in IR (Goff and Horst, 1997;
Rukkwamsuk et al., 2000; Bobe et al., 2004; Leroy et al., 2005; Pires et at., 2007; Bossaert et al., 2008c;
Schoenberg en Overton, 2010).
Door de perifere IR en de chronische lage insuline spiegels ziet men ook dat bepaalde ziekten zoals de
ketonemie en de leververvetting de neiging hebben om steeds te verergeren (Van Winden et al., 2003a
en b; Hayirli, 2006; Bossaert et al., 2008a).
Ziekten op zich hebben ook een invloed op de IR, zo ziet men bij leververvetting en ketoacidose een
toegenomen IR (Hayirli, 2006; Pires et al., 2007; Bossaert et al., 2008a).
Insuline secretie
Soms ziet men, zoals men ook al bij de mens gezien heeft dat bij hoogproductief melkvee de insuline
secrectie door de pancreas sterk aangetast is. Zo werden er tijdens studies over koeien met cysteuze

28
ovariële follikels van Opsomer et al. (1999) en over koeien met ketonemie van Hove (1978) een aantal
koeien ontdekt die niet reageerden met een toename van insuline secretie na het toedienen van een
intraveneuze glucose bolus. Deze verminderde insuline secretie t.h.v. de pancreas is o.a. te wijten aan de
hoge gehaltes van NEFA’s in de periode rond de geboorte (Bossaert et al., 2008c). Hieruit kan er besloten
worden dat er sommige koeien, net zoals sommige mensen gevoelige β-cellen hebben die onderdrukt
kunnen worden door de chronische verhoging van NEFA’s (Jensen, 2006; Kahn, 2006; De Koster en
Opsomer, 2012). Deze veranderingen van insuline secretie door de β-cellen werden ook al gevonden bij
neonatale kalveren waardoor er een groot vermoeden is dat dit (epi)genetisch bepaald is (Bossaert et al,
2009).
3.2.3. Glucagon
Glucagon is eigenlijk de antagonist van insuline (Samuelson Don, 2007). De belangrijkste stimulus voor
de vrijstelling van het hormoon is een verlaagde glucosespiegel (Frandson et al., 2003). In de lever
stimuleert glucagon de glycogenolyse en de gluconeogenese, waardoor er veel glucose vrijkomt en er een
verhoging is van de bloedglucosespiegel (Brockman en Laarveld, 1986; Frandson et. al., 2003). In het
vetweefsel wordt de lipolyse gestimuleerd, nl. door stimulatie van het enzym hormoon sensitive lipase,
waardoor er NEFA’s zullen vrijkomen (Frandson et. al., 2003; Bossaert et al., 2008b).
Tijdens de lactatie is de belangrijkste taak van glucagon de stimulatie van gluconeogenese t.h.v. de lever
en blijkt de werking ter hoogte van de perifere weefsels eerder beperkt te zijn. Glucagon blijkt
voornamelijk belangrijk te zijn in stress-situaties (Hart, 1983; Opsomer, 1995).
Ter hoogte van .de lever blijkt voornamelijk de verhouding insuline/glucagon belangrijk te zijn daar deze
een antagonistische werking hebben. Hoe lager deze verhouding, hoe groter de hoeveelheid glucose die
ter beschikking kan gesteld worden voor de lactatie, en dit zou ook gepaard gaan met een verhoogde
ketonemie (Hart, 1983; Herdt, 1988; Torck, 1993; Opsomer 1995). Gedurende de ganse lactatie blijkt de
glucagonconcentratie vrij stabiel te zijn en is de insuline/glucagon verhouding dus voornamelijk afhankelijk
van de schommelingen in de insuline concentratie. In het begin van de lactatie is het insuline zeer laag
waardoor de glucagon-effecten dan de overhand hebben en tegen het einde van de lactatie stijgt de
concentratie van insuline terug waardoor de insuline-effecten, nl. opslaan van reserves dan de bovenhand
hebben en de reserves aangevuld worden voor de volgende lactatie (Opsomer, 1995; Bossaert et al.,
2008b).
Glucagon zou ook kunnen gebruikt worden in de preventie van leververvetting en ketonemie, maar dit is
echter enkel theoretisch daar glucagon niet mag gebruikt worden bij melkvee (Hayirli et al., 2002; Nafikov
et al., 2002; Bobe et al., 2004; Cools et al., 2008b).
3.2.4. Pancreas polypeptide
De specifieke rol van pancreas polypeptide (PP) is nog niet volledig opgehelderd maar het zou een rol
spelen in de vrijstelling van de verteringssappen en de voederopname en zou dus indirect invloed hebben
op het glucosegehalte in het bloed. De vrijstelling van PP staat onder vagale controle en blijkt beïnvloed
door de entero-pancreatische reflex. Inname van eiwitten, vasten, arbeid en acute hypoglycemie
stimuleren de secretie van PP (Martin en Crump, 2003).

29
3.2.5. Somatostatine Pancreas polypeptide
Somatostatine, of ook soms somatotropine release inhibiting factor (SRIF) genoemd, wordt geproduceerd
door vele verschillende weefsel waaronder de hypothalamus, de pancreas (door de delta-cellen) en de
maag. De vrijstelling van somatostatine wordt gestimuleerd door glucose, aminozuren, vetzuren,
gastrointestinale hormonen, glucagon en bèta-adrenerge neurotransmittors. De vrijstelling wordt
geïnhibeerd door insuline en alfa-adrenerge neurotransmittors (Martin en Crump, 2003).
Somatostatine remt op zijn beurt dan weer o.a. de vrijstelling van glucagon, insuline en groeihormoon en
pancreassap. Ze verlaagt ook de motiliteit van het maagdarmkanaal en de galblaas. De werking van
somatostatine bij de melkproductie is dus niet rechtstreeks maar vnl. door invloed op de andere hormonen
insuline, glucagon en groeihormoon (Martin en Crump, 2003; Reimers, 2003; Samuelson Don, 2007).
3.2.6. Insuline amyloid polypeptide (IAPP) of amylin
Insuline amyloïd polypeptide (IAPP), ontstaat uit pro-islet amyloid polypeptide (proIAPP), is samen
opgeslagen en vrijgesteld met insuline.
De specifieke functies zijn nog niet exact gekend bij de herkauwers, maar in de humane geneeskunde is
hier wel al meer over geweten. Insuline amyloid polypeptide heeft de mogelijkheid om de insuline
gestimuleerde glycogenesis in de spieren te inhiberen, het schaadt de effecten van insuline op de glucose
output van de lever en vermindert de opname van glucose door perifere weefsels. Het heeft ook een
calcitonine-achtige werking in het botmetabolisme en een calcitonin gene related peptide (CGRP)-achtige
werking op de vascularisatie. Ook zou het een rol spelen in de voedselopname. Men ziet ook dat IAPP
een rol speelt in het ontstaan van diabetes type 2, nl. door het induceren van apoptose in insuline
producerende bèta-cellen door het neerslaan van amyloïd, gevolgd door een verminderde insuline
secretie (Lorenzo et al., 1994; Pittner et al., 1994; Kahn, 2003; Martin en Crump, 2003; Paulsson en
Westermark, 2005; Paulsson et al., 2006).

30
BESPREKING
De waarneming dat de sterk toegenomen melkproductie gepaard gaat met een verhoogde incidentie van
metabole problemen dwingt ons om de onderliggende oorzaken hiervan te onderzoeken (van Arendonk
en Liinamo, 2003; Opsomer et al., 2006; Bossaert et al., 2008a).
Deze invloeden op productie en gezondheid zijn zeer nauw met elkaar verbonden, waardoor het niet
steeds mogelijk is om een scheidingslijn te trekken tussen het “fysiologische” en het “pathologische”. Zo
komen hoogproductieve dieren bij het begin van de lactatie vaak in een negatieve energiebalans terecht
en vindt er een uitgebreide lipolyse plaats. Dit is tot op een bepaald niveau nuttig en kan als normaal
aanzien worden, daar de vrijgekomen stoffen kunnen gebruikt worden als energieleveranciers. Echter
wanneer deze lipolyse te erg wordt, kunnen er zich metabole ziekten, zoals leververvetting en ketonemie
ontwikkelen. Deze ziekten zorgen op hun beurt voor een verhoogd risico op het ontstaan van andere
ziekten en een daling van de productie, waardoor de dieren dus in een negatieve spiraal kunnen
terechtkomen. De metabole ziekten hebben vaak een vaag en aspecifiek symptomenbeeld en liggen in
het verlengde van de normale adaptatiemechanismen (Veenhuizen et al., 1991; Jorritsma et al., 2003;
Bobe et al., 2004; Bossaert et al., 2008a; Schoenberg en Overton, 2010).
Ondanks deze moeilijkheid is men er toch al in geslaagd om verschillende linken bloot te leggen. Bekend
is bijvoorbeeld dat de endocriene pancreas een belangrijke regulerende rol heeft bij het hoogproductieve
melkvee. Hierbij is de rol van insuline al goed onderzocht en zijn er al enkele belangrijke ontdekkingen
gedaan zoals bv. de verschillen in glucose receptoren (GLUT-typen) ter hoogte van de verschillende
weefsels wat ervoor zorgt dat er een specifieke verdelingvan glucose kan plaatsvinden (Hayirli, 2006;
Bossaert, 2010; Schoenberg en Overton, 2010).Toch zijn er ook enkele opmerkelijke bevindingen en
soms tegenstrijdigheden te vinden. Zo spelen bijvoorbeeld bij een lebmaagverplaatsing zowel een
verhoogde als een verlaagde insuline concentratie een rol (Van Winden et al., 2003a; Doll et al., 2009;
Türck en Leonhard-Marek, 2010).
Hierbij bestaat dus de opdracht om nog beter de werking en regeling van insuline, en de daarbij horende
insuline secretie t.h.v. de pancreas en insuline resistentie t.h.v. de perifere weefsels te onderzoeken, om
dan preventief te kunnen ingrijpen op het ontstaan van metabole ziekten en een hoge productie te
waarborgen.Hierbij is er vnl. nog verder onderzoek nodig naar de factoren die de insuline secretie, en
meer specifiek de β-cellen bij de melkkoe beïnvloeden. Zo is er een sterk vermoeden dat ook
epigenetische factoren een rol spelen (Bossaert, 2009; De Koster en Opsomer, 2012).
Naast de rol van insuline en glucagon als antagonist waar al veel over is gekend is, is er ook nog verder
onderzoek nodig naar de functies en werking van de andere pancreashormonen zoals somatostatine,
pancreaspolypeptide (PP) en insuline amyloid polypeptide (IAPP) bij het hoogproductief melkvee.
Hiervoor kan men zich ook baseren op de rijkere kennis hieromtrent in de humane geneeskunde. Zo is bv.
in de humane geneeskunde ontdekt dat IAPP o.a. aanleiding geeft tot neerslaan van amyloïd en apoptose
van de β-cellen, wat eveneens zal leiden tot een verminderde insuline secretie (Lorenzo et al., 1994;
Pittner et al., 1994; Kahn, 2003; Martin en Crump, 2003; Paulsson en Westermark, 2005; Paulsson et al.,
2006, Kahn, 2003; Bossaert, 2009; De Koster en Opsomer, 2012)
Om de ziekten beter te kunnen aanpakken in de praktijk wil men metabole parameters bepalen, die men
al zou kunnen bepalen alvorens er symptomen zijn en om zo preventief te kunnen ingrijpen. Dit is echter
niet steeds even eenvoudig daar de etiologie en pathogenese van vele ziekten nog niet volledig ontrafeld
zijn. De resultaten van verschillende studies i.v.m. deze parameters spreken elkaar nog vaak tegen

31
waardoor ze in de praktijk nog niet echt bruikbaar zijn. Dit vormt dus een grote uitdaging om deze
parameters in de toekomst te vinden (Bobe et al., 2004; Opsomer et al., 2006; Bossaert et al., 2008a;
Cools et al., 2008b).

32
REFERENTIELIJST
1. Akers R.M. en Denbow D.M. (2008). Anatomy & physiology of domestic animals, First edition.
Blackwell Publishing, Ames, Iowa, p. 475-500.
2. Baird G.D. (1982). Primary ketosis in the high-producing dairy cows: clinical and subclinical
disorders, treatment, prevention, and outlook. Journal of Dairy Science 65, 1-10.
3. Barone R. (2009). Splanchnologie I. In: Anatomie compare des mammifières domestiques, Tome 3.
Vigotéditeurs, Paris, p. 472, p. 570-571.
4. Bauman D.E., Currie W.B. (1980). Partitioning of nutrients during pregnancy and lactation: a review
of mechanisms involving homeostasis and homeorhesis. Journal of Dairy Science 63 (9), 1514-1529.
5. Beever D.E. (2006). The impact of controlled nutrition during the dry period on dairy cow health,
fertility and performance. Animal Reproduction Science 96, 212-226.
6. Bell A.W. (1995). Regulation of organic nutrient metabolism during transition from late pregnancy to
early lactation. Journal of Animal Science 73, 2804-2819.
7. Bell W.A. and Bauman D.E. (1997). Adaptation of glucose metabolism during pregnancy and
lactation. Journal of Mammary Gland Biology and Neoplasia, vol.2, no.3, 265-278.
8. Berne R.M, Koeppen B. M., Bruce A. S. (2010). Hormones of the pancratic islets. Berne & Levy
physiology, 6th edition, Mosby/Elsevier, Philadelphia, 836pgs.
9. Berry D.P., Buckley F., Dillon P., Evans R.D., Rath M., Veerkamp R. F. (2002). Genetic parameters
for level and change of body condition score and body weight in dairy cows. Journal of Dairy Science
85, 2030-2039.
10. Bertics S.J., Grummer R.R., Cadorniga-Valino C., Stoddard E.E. (1992). Effect of prepartum dry
matter intake on liver triglyceride concentration and early lactation. Journal of Dairy Science 75,
1914-1922.
11. Bines J. A. (1976). Regulation of food intake in dairy cows in relation to milk production. Livestock
Production Science 65, 1375-1389.
12. Bobe G., Young J.W., Beitz D.C. (2004). Invited Review: Pathology, etiology, prevention, and
treatment of fatty liver in dairy cows. Journal of Dairy Science 87, 3105-3124.
13. Bonner-Weir S., Like A.A. (1980). A dual population of islets of Langerhans in bovine pancreas.
Celland Tissue Research volume 206, number 1, 157-170.
14. Bossaert P. (2010). The role of insulin in the energy conflict between milk production and ovarian
activity during the transition period of high-yielding dairy cows. Proefschrift voorgedragen tot het
behalen van de graad van Doctor in de Diergeneeskundige Wetenschappen (PhD) aan de Faculteit
Diergeneeskunde, Universiteit Gent, 248pgs.
15. Bossaert P., Cools S., Van Loo H., Leroy J., de Kruif A., Opsomer G. (2008a). De pathogenese en de
kliniek van ketonemie en leververvetting bij hoogproductieve melkkoeien. Vlaams Diergeneeskundig
Tijdschrift 77, 283-289.
16. Bossaert P., Leroy J., Cools S., Van Loo H., de Kruif A., Opsomer G. (2008b). De metabole
adaptatiemechanismen bij hoogproductieve melkkoeien. Vlaams Diergeneeskundig Tijdschrift 77,
207-215.
17. Bossaert P., Leroy J., De Vlieger S., Opsomer G. (2008c). Interrelations between glucose-induced
Insulin response, metabolic indicators and time of first ovulation in High Yielding Dairy Cows. Journal
of Dairy Science 91, 3363–3371.

33
18. Bossaert P, Leroy J.L.M.R., De Campeneere S.,De Vliegher S., Opsomer G. (2009). Differences in
the glucose-induces insulin response and the peripheral insulin responsiveness between neonatal
calves of the Belgium Blue, Holstein-Friesian and East Flemish breeds. Journal of Dairy Science 92,
4401-4411.
19. Braw-Tal R., Pen S., Roth Z. (2009). Ovarian cysts in high-yielding dairy cows. Theriogenology 72,
690–698.
20. Brockman R.P. en Laarveld B. (1986). Hormonal regulation of metabolism in ruminants: a review.
Livestock Production Science 14, 313-334.
21. Butler S.T., Pelton S.H., Butler W.R.(2004). Insulin increases 17β-estradiol production by the
dominant follicle of the first postpartum follicle wave in dairy cows. Reproduction 127, 537-545.
22. Chagas L.M., Lucy M.C., Back P.J., Blache D., Lee J.M., Gore P.J.S., Sheahan A.J., en Roche J.R.
(2009). Insulin resistance in divergent strains of Holstein-Friesian dairy cows offered fresh pasture
and increasing amounts of concentrate in early lactation. Journal of Dairy Science 92, 216–222.
23. Christensen J.O., Grummer R.R., Rasmussen F.E. Bertics S.J. (1997). Effect of Method of Delivery of
Propylene Glycol on Plasma Metabolites of Feed-Restricted Cattle. Journal of Dairy Science 80, 563-
568.
24. Coffey M.P., Simm G., Oldham J.D., Hill W.G., Brotherstone S. (2004). Genotype and diet effects on
energy balance in the first three lactations of dairy cows. Journal of Dairy Science 87, 4318-4326.
25. Cools S., Bossaert P., Caluwaerts, Hostens M., Opsomer G., de Kruif A. (2008a). De economische
gevolgen van een verlenging van de tussenkalftijd bij hoogproductief melkvee. Vlaams
Diergeneeskundig Tijdschrift 77, 406-413.
26. Cools S., Bossaert P., Van Loo H., de Kruif A., Opsomer G. (2008b). De behandeling en preventie
van ketonemie en leververvetting bij hoogproductieve melkkoeien. Vlaams Diergeneeskundig
Tijdschrift 77, 363-375.
27. Coppock C.E. (1985). Energy nutrition and metabolism of the lactating dairy cow. Journal Dairy
Science 68, 3403-3410.
28. Coppock. C.E., Everett R.W., Natzke R.P. (1974). Effect of dry period length on holstein milk
production and selected disorders at parturition. Journal Dairy Science 57, 712-718.
29. Cornier M., Dabelae D., Hernandez T.L., Lindstrom R.C., Steig A.J., Stob N.R., Van Pelt R.E., Wang
H., Eckel R.H. (2008). The metabolic syndrome. Endocrine Reviews 29, 777-822.
30. Cunningham J.G., Klein B. G. (2007). Textbook of veterinary physiology, 4th edition. Saunders, St-
Louis, Missouri, p. 501-516.
31. De Koster J. en Opsomer G. (2012). Are modern dairy cows suffering from modern diseases?.
Vlaams Diergeneeskundig Tijdschrift 81, 71-80.
32. de Kruif A. (2008). Tussenkalftijd blijft toenemen. Melkveebedrijf nr.3, 16-17.
33. Deprez P. (2010). Lesnota’s bij de cursus pathofysiologie en klinische chemie. Cursus faculteit
Diergeneeskunde, Gent, 90pgs.
34. Deprez P. en van Loon G. (2012). Inwendige ziekten van de grote huisdieren, Cursus faculteit
Diergeneeskunde, Gent, 279pgs.
35. Doll K., Sickinger M., Seeger T. (2009). New aspects in the pathogenesis of abomasal displacement.
The Veterinary Journal 181, 90-96.
36. Drackley, J.K. (1999). Biology of dairy cows during the transition period: The final frontier? Journal
DairyScience 82, 2259–2273.

34
37. Drackley J.K., Dann H.M., Douglas G.N., Guretzky N.A.J., Litherland N.B., Underwood J.P., Loor J.J.
(2005). Physiological and pathological adaptations in dairy cows that may increase susceptibility to
periparturient diseases and disorders; Italian Journal of Animal Science 4, 323-344.
38. Edmonson A.J., Lean I.J., Weaver L.D., Farver T., Wester G.(1989). A body condition scoring chart
for Holstein dairy cows. Journal Dairy Science 72, 68-78.
39. Ellenberger W., Baum H., Zietzschmann O. (1974). Handbuch der vergleichenden Anatomie der
Haustiere. 18e Auflage. Springer, Berlin, Heidenberg, New York, p. 451.
40. Frandson R.D., Wile W.L., Fails A.D. (2003). Anatomy and physiology of farm animals, 6th edition.
Lea and Febiger, Philadelphia, p. 415-427.
41. Garnsworthy P.C. and Topps J.H. (1982). The effect of body condition of dairy cows at calving on
their food intake and performance when given complete diets. Animal Production 35, 113-119.
42. Goff J.P., Horst R.L. (1997). Physiological changes at parturition and their relationship to metabolic
disorders. Journal of Dairy Science 80, 1260-1268.
43. Grummer R.R. (1993). Etiology of lipid-related metabolic disorders in periparturient dairy cows.
Journal of Dairy Science 76, 3882-3896.
44. Grummer R.R., Mashek D.G., Hayirli A. (2004). Dry matter intake and energy balans in the transition
period. Veterinary Clinics of North America, Food Animal Practice 20, 447-470.
45. Hart I.C. (1983). Endocrine control of nutrient partition in lactating ruminants. Proceedings of the
Nutrition Society 42, 181-194.
46. Hayirli A. (2006). The role of exogenous insulin in the complex of hepatic lipidosis and ketosis
associated with insulin resistance phenomenon in postpartum dairy cattle. Veterinary Research
Communications 30(7), 749-774.
47. Hayirli A, Bertics S.J., Grummer R.R. (2002). Effects of slow-release insulin on production, liver
triglyceride, and metabolic profiles of Holsteins in early lactation. Journal of Dairy Science 85, 2180-
2191.
48. Herdt T. H. (1988). Fuel homeostasis in the ruminant. Vet. Clin. North Am., Food Animal Practice,
vol. 4, no 2, 213-231.
49. Herdt T.H. (2000). Ruminant adaptation to negative energy balance. Influences on the etiology of
ketosis and fatty liver. The Veterinary Clinics of North America. Food Animal Practice 16, 215-230.
50. Hermans K. (2010). Metabool profiel als hulpmiddel om de transitieperiode van hoogproductief
melkvee te beoordelen. Onderzoeksverslag in kader van de masterproef Faculteit diergeneeskunde,
Gent, 76pgs.
51. Heuer C., Schukken Y.H., Dobbelaar P. (1999). Postpartum body condition score and results from
the First test day milk as predictors of disease, fertility, yield, and culling in commercial dairy herds.
Journal of Dairy Science 82, 9. 295-304.
52. Hippen A.R., She P., Young J.W., Beitz D.C., Lindberg G.L., Richardson L.F., Tucker R.W. (1999).
Alleviation of fatty liver in dairy cows by 14-days intravenous infusions of glucagon. Journal of Dairy
Science 82, 1139-1152.
53. Holtenius P.en Holtenius K. (1996): New aspects of ketone bodies in energy metabolism in dairy
cows: A Review. Zentralbl Veterinarmed A. 43, 579-587.
54. Holtenius K., Agenäs S., Delavaud C., Chilliard Y. (2003). Effects of feeding intensity during the dry
period. 2. Metabolic and hormonal responses. Journal of Dairy Science 86, 883-891.
55. Hostens M. (2008). Bestaat het ideale droogstand rantsoen? Melkveebedrijf nr.4, 18-20.

35
56. Hove K. (1978). Insulin secretion in lactating cows: responses to glucose infused intravenously in
normal, ketonemic and starved animals. Journal of Dairy Science 61, 1407-1413.
57. Janssens G. en Hesta M. (2011). Dierenvoeding. Cursus faculteit Diergeneeskunde, Gent, 281pgs.
58. Jensen M.D. (2006). Adipose tissue as an endocrine organ: implications of its distribution on free
fatty acid metabolism. European Heart Journal Supplements 8, 13-19.
59. Johannsen U., Menger S., Staufbiel R., Klukas H. (1993). The morfology and function of the liver in
high performance cows 2 weeks post partum. Deutsche TierarztlWochenschr 100(5), 177-181 (in
German, abstract in English).
60. Jorritsma R., Jorritsma H., Schukken Y.H., Bartlett P.C., Wensing T., Wentink G.H. (2001).
Prevalence and indicators of post partum fatty infiltration of the liver in nine commercial dairy herds in
the Netherlands. Livestock Production Science 68, 53-60.
61. Jorritsma R., Wensing T., Kruip T.A.M., Vos P.L.A.M., Noordhuizen J.P.T.M. (2003). Metabolic
changes in early lactation and impaired reproductive performance in dairy cows. Veterinary Research
34,11-26.
62. Jorritsma R., Jorritsma H., Schukken Y.H., Wentink G.H. (2000). Relationships between fatty liver
and fertility and some periparturient diseases in commercial Dutch dairy herds. Theriogenology 54,
1065-1074.
63. Kahn C.R. (1978). Insulin resistance, insulin sensitivity and insulin unresponsiveness: a necessary
distinction. Metabolism 27, 1893-1902.
64. Kahn S.E. (2003). Review: the relative contributions of insulin resistance and beta-cell dysfunction to
the pathophysiology of type 2 diabetes. Diabetologia 46 , 3-19.
65. Kahn S.E., Hull R.L., Utzschneider K.M. (2006). Mechanisms linking obesity to insulin resistance and
type 2 diabetes. Nature 444, 840-846.
66. Katoh N. (2002). Relevance of apolipoproteins in the development of fatty liver and fatty liver-related
peripartum diseases in dairy cows. Journal of Veterinary Medical Science 64, 293-307.
67. Kawashima C., Fukihara S., Maeda M., Kaneko E., Montoya C.A., Matsui M., Shimizu T, Matsunaga
N, Kida K, Miyake Y, Schams D, Miyamoto A. (2007). Relationship between metabolic hormones and
ovulation of dominant follicle during the first follicular wave post-partum in high-producing dairy
cows.Reproduction 133, 155–163.
68. Kerestes A.M. (2009). Role of insulin in the development of metabolic and reproductive malfunctions
of periparturient dairy cows. PhD, SzentIstván university, faculty of veterinary science, Budapest,
116pgs.
69. Kim I.H., Suh G.H. (2003). Effect of the amount of body condition loss from the dry to near calving
periods on the subsequent body condition change, occurrence of postpartum diseases, metabolic
parameters and reproductive performance in Holstein dairy cows. Theriogenology 60, 1445-1456.
70. Kleen J.L., Stokman P., Noordhuizen J.P.T.M., Rehage J., Hooijer G. (2003). Subacuteruminal
acidosis (SARA) in dairy cows. ‘From research to clinic’. In: Proceedings of the Journées
Européennes de Buiatrie, Paris, 24-30.
71. Knight C.H., France J., Beever D.E. (1994). Nutrient metabolism and utilization in the mammary
gland. Livestock Production Science 39, 129-137.
72. Kokkonen T. (2005). Energy and protein nutrition of dairy cows during the dry period and early
lactation: production performance and adaptation from pregnancy to lactation. Academic Dissertation,
Faculty of Agriculture and Forestry, University of Helsinki, 66pgs.

36
73. Kräft S. (2004). Charakterisierung der peripheren Insulin-Response und Insulin-Sensitivität bei
trockenstehenden, laktierenden und leberverfetteten Milckkühen ohne und mit Ketose mittels
hyperinsulinämischer, euglycämischer Clamps. Academic Dissertation Tierärztliche Hockschule
Hannover, 253pgs.
74. Leblanc S.J., Leslie K.E., Duffield T.F. (2005). Metabolic predictors of displaced abomasum in dairy
cattle. Journal of Dairy Science 88, 159-170.
75. Leroy J.L.M.R., Vanholder T., Mateusen B., Christophe A., Opsomer G., de Kruif A., Genicot G., Van
Soom A. (2005). Non-esterified fatty acids in follicular fluid of dairy cows and their effect on
developmental capacity of bovine oocytes in vitro. Reproduction 130, 485-495.
76. Lorenzo A, Razzaboni B, Weir GC, Yankner BA (1994). Pancreatic islet cell toxicity of amylin
associated with type-2 diabetes mellitus. Nature 368, 756–760.
77. Marcos E., Mazur A., Cardot P, Rayssiguier Y. (1990). Serum apolipoproteins B and A-I and naturally
occurring fatty liver in dairy cows. Lipids 25, 575-577.
78. Martin P.A. en Crump M.H. (2003). The endocrine pancreas. In: Mc Donald L. E., Pineda M. H. en
Dooley M. P. (ed), Veterinary Endocrinology and Reproduction, 5th edition, Iowa State Press, Ames,
Iowa, p. 141-163.
79. McDowell G.H. (1991). Somatotropin and endocrine regulation of metabolism during lactation.
Journal of Dairy Science 74, 44-62.
80. McNamara J.P., Hillers J.K. (1986). Adaptations in lipid metabolism of bovine adipose tissue in
lactogenesis and lactation. Journal of Lipid Research 27, 150-157.
81. Moe P.W. (1981). Energy metabolism of dairy cattle. Journal of Dairy Science, 64, p1120-1139.
82. Morrow D. A. (1976). Fat cow syndrome. Journal Dairy Science 59, 1625-1629.
83. Murondoti A., Jorritsma R., Beynen A.C., Wensing T., Geelen M.J. (2004). Activities of the enzymes
of hepatic gluconeogenesis in periparturient dairy cows with induced fatty liver. Journal of Dairy
Science 71, 129-134.
84. Nafikov R.A., Ametaj B.N., Bobe G., Koehler K.J., Young J.W., Beitz D.C. (2002). Prevention of fatty
liver in transition dairy cows by subcutaneous injections of glucagon. Journal of Dairy Science 89,
1533-1545.
85. Nickel R., Schummer A., Sack W.O. (1979). The viscera of domestic animals. 2nd rev. edition. Parey,
Berlin, p. 119-122 en p. 179-180.
86. Nocek J.E. en Braund D.G. (1985). Effect of feeding frequency on diurnal dry matter and water
consumption, liquid dilution rate and milk yield in first lactation. Journal of Dairy Science 68, 2238-
2247.
87. Opsomer G. (1995). Het energiemetabolisme bij hoogproductief melkvee: een literatuurstudie.
Eindstudiewerk ingediend tot het behalen van het diploma van gespecialiseerde in de dierlijke
productie, Faculteit diergeneeskunde, Universiteit Gent, 86 pgs.
88. Opsomer G., Coryn M., Deluyker H., de Kruif A. (1998). An analysis of ovarian dysfunction in high
yielding dairy cows after calving based on progesterone profiles. Reproduction in Domestic Animals
33, 193-204.
89. Opsomer G., de Kruif A. (1998). De behandeling en preventie van hypocalcemie of kalfziekte bij
hoogproduktief melkvee: een praktische benadering. Vlaams Diergeneeskundig Tijdschrift 67, 310-
314.

37
90. Opsomer G., De Vliegher S., de Kruif A. (2004). Hoe kan het droogzetten van hoogproductieve
melkkoeien in de praktijk het best gebeuren? Vlaams Diergeneeskundig Tijdschrift 73, 384-395.
91. Opsomer G., Gröhn Y., Hertl J., Coryn M., Deluyker H., de Kruif A. (2000). Risk factors for post
partum ovarian dysfunction in high producing dairy cows in Belgium: a field study. Theriogenology 53,
841-857.
92. Opsomer G., Leroy L., Vanholder T., Bossaert P., de Kruif A. (2006). Subfertility in high yielding dairy
cows: how to bring science into practice? Vlaams Diergeneeskundig Tijdschrift 75, 114-120.
93. Opsomer G., Wensing Th., Laevens H., Coryn M., de Kruif A. (1999). Insulin resistance: link between
metabolic disorders and cystic ovarian disease in high yielding dairy cows?. Animal Reproduction
Science 56, 211-222.
94. Paulsson J.F., Andersson A., Westermark P., Westermark G.T. (2006). Intracellular amyloid-like
deposits contain unprocessed pro-islet amyloid polypeptide (proIAPP) in beta cells of transgenic mice
overexpressing the gene for human IAPP and transplanted human islets. Diabetologia 49, 1237–
1246.
95. Paulsson J.F., Westermark G.T. (2005). Aberrant processing of human proislet amyloid polypeptide
results in increased amyloid formation. Diabetes 54, 2117–2125.
96. Picket M.M., Piepenbrink M.S., Overton T.R (2003). Effects of propylene glycol or fat drench on
plasma metabolites, liver composition, and production of dairy cows during the periparturient period.
Journal of DairyScience 86, 2113-2121.
97. Pires J.A.A., Souza A.H., Grummer R.R. (2007). Induction of Hyperlipidemia by Intravenous Infusion
of Tallow Emulsion Causes Insulin Resistance in Holstein Cows. Journal of Dairy Science 90, 2735-
2744.
98. Pittner R.A., Albrandt K., Beaumont K., Gaeta L.S., Koda J.E., Moore C.X., Rittenhouse J., Rink T.J.
(1994). Molecular physiology of amylin. Journal of Cellular Biochemistry 55, 19–28.
99. Pryce J. E., Royal M.D., Garnsworthy P.C., and Mao. I.L. (2004). Fertility in the high-producing dairy
cow. Livestock Production Science 86, 125-135.
100. Rehage J., Qualmann K., Meier C., Stockhofe-Zurweiden N., Holtershinken M., Pohlenz J. (1999).
Total serum bile acid concentrations in dairy cows with fatty liver and liver failure. Deutsche Tierarztl
Wochenschr. 106(1), 26-29. (in German, abstract in English)
101. Reimers T.J. (2003). The Pituitary gland. In: Mc Donald L. E., Pineda M. H. en Dooley M. P. (ed),
Veterinary Endocrinology and Reproduction, 5th edition, Iowa State Press, Ames, Iowa, p. 17-34.
102. Roche J.R., Friggens N.C., Kay J.K., Fisher M.W., Stafford K.J., Berry D.P. (2009). Invited review:
Body condition score and its association with dairy cow productivity, health, and welfare. Journal of
Dairy Science 92, 5769-5801.
103. Rukkwamsuk T., Geelen M.J. H., Kruip T.A.M., Wensing T. (2000). Interrelation of fatty acid
composition in adipose tissue, serum and liver of dairy cows during the development of fatty liver
postpartum. Journal of Dairy Science 83, 52-59.
104. Rukkwamsuk T., Kruip T.A.M., Wensing T. (1999a). Relationship between overfeeding and
overconditioning in the dry period and the problems of high producing dairy cows during the
postparturient period. The Veterinary Quarterly 21, 71-77.
105. Rukkwamsuk T., Wensing T., Geelen M.J.H (1998). Effect of overfeeding during the dry period on
regulation of adipose tissue metabolism in dairy cows during the periparturient period. Journal of
Dairy Science 81, 2904-2911.

38
106. Rukkwamsuk T., Wensing T., Geelen M.J.H. (1999b). Effect of fatty liver on hepatic
gluconeogenesis in periparturient dairy cows. Journal of Dairy Science 82, 500-505.
107. Rukkwamsuk T., Wensing T., Geelen M.J.H. (1999c). Effect of overfeeding during the dry period
on the rate of esterification in adipose tissue of dairy cows during the periparturient period. Journal of
Dairy Science 82, 1165-1169.
108. Samuelson Don A. (2007). Textbook of veterinary histology. Saunders-Elsevier, St. Louis,
Missouri, p. 355-358 en p. 416.
109. Sano H., Nakai M., Kondo T., Terashima Y. (1991). Insulin responsiveness to glucose and tissue
responsiveness to insulin in lactating, pregnant, and nonpregnant, nonlactating beef cows. Journal of
Animal Science 69, 1122-1127.
110. Saul M. G. (2000). Endocrine system. In: Berne R. and Levy M. (editors) Principles of physiology,
third edition, Mosby, St-louis, Missouri, p. 50-547.
111. Scalia D., Lacetera N., Bernabucci U., Demeyere K., Duchateau L., Burvenich C. (2006). In vitro
effects of nonesterified fatty acids on bovine neutrophils oxidative burst and viability. Journal of Dairy
Science 89, 147-154.
112. Schoenberg K.M. en Overton T.R. (2010). The changing roles of insulin during the transition
period. Proceedings, Cornell Nutrition Conference For Feed Manufacturers,175-185
113. Silbernagl S. en Despopoulos A. (2008). Sesam atlas van de fysiologie. HBuitgevers, Baarn
p.228-311.
114. Spicer L.J., Alpizar E., Echternkamp S.E. (1993). Effects of insulin, insulin-like growth factor I, and
gonadotrophins on bovine granulosa cell proliferation, progesterone production, estradiolproduction,
and (or) insulin-like growth factor I production in vitro. Journal of Animal Science 71, 1232–1241.
115. Spicer L.J., Echternkamp S.E. (1995). The ovarian insulin and insulin-like growth factor system
with an emphasis on domestic animals. Domestic Animal Endocrinology 12, 223-245.
116. Strang B.D., Bertics S.J., Grummer R.R., Armentano L.E. (1998). Relationship of triglyceride
accumulation to insulin clearance and hormonal responsiveness in bovine hepatocytes. Journal of
Dairy Science 81, 740-747.
117. Studer V.A., Grummer R.R., Bertics S.J., Reynolds C.K. (1993). Effect of prepartum propylene
glycol administration on periparturient fatty liver in dairy cows. Journal of Dairy Science 76, 2931-
2939.
118. Suriyasathaporn W., Heuer C., Noordhuizen-Stassen E.N., Schukken Y.H. (2000). Hyper-
ketonemia and the impairment of udder defense: a review. Veterinary Research 31, 397-412.
119. Swali A. en Wathes D.C. (2006). Influence of the dam and sire on size at birth and subsequent
growth, milk production and fertility in dairy heifers. Theriogenology 66,1173–1184.
120. Torck T. (1993). Metabole profielen bij hoogproductief melkvee: factoren die het normale verloop
van de belangrijkste energieparameters beïnvloeden. Eindstudiewerk, Faculteit diergeneeskunde,
Universiteit gent, 69pgs.
121. Treacher R.J., Reid I.M. and Roberts C.J. (1986). Effect of body condition at calving on the health
and performance of dairy cows. Animal Production 43, 1-6.
122. Türck G. en Leonhard-Marek S. (2010). Potassium and insulin affect the contractility of abomasal
smooth muscle. Journal of Dairy Science 93, 3561-3568.
123. Van Arendonk J.A.M. , Liinamo A. (2003). Dairy cattle production in Europe. Theriogenology 59,
563-569.

39
124. Van der Putten, M. (2005). Insulinewerking bij hoogproductief melkvee, met name in de
postpartum periode, scriptie voorgedragen tot het behalen van het diploma van dierenarts, Faculteit
diergeneeskunde, Universiteit Gent, 34pgs.
125. Van der walt J.G. en Linington M.J. (1989). A review of energy metabolism in producing
ruminants. Part 1: Metabolism of energy substrates. J. South Afr. Vet. Ass. 60, 223-227.
126. Vanholder T., Leroy J.L.M.R., Dewulf J., Duchateau L., Coryn M., de Kruif A., OpsomerG. (2005).
Hormonal and metabolic profiles of high-yielding dairy cows prior to ovarian cyst formation or first
ovulation post partum. Reproductionin Domestic Animals, 40, 460–467.
127. Vanholder T., Leroy J.L.M.R., Van Soom A., Opsomer G., Maes D., Coryn M., de Kruif A. (2005).
Effect of non-esterified fatty acids on bovine granulosa cell steroidogenesis and proliferation in vitro.
Animal Reproduction Science 87, 33-44.
128. Van Soest P.J. (1994). Nutritional ecology of the ruminant. Cornell University Press, 2nd edition,
476pgs.
129. Van Winden S.C.L., Jorritsma R., Muller K.E., Noordhuizen J.P.T.M. (2003 a). Feed intake, milk
yield, and metabolic parameters prior to left displaced abomasum in dairy cows. Journal of Dairy
Science 86, 1465-1471.
130. Van Winden S.C.L., Kuiper R. (2003 b). Left displacement of the abomasum in dairy cattle: recent
developments in epidemiological and etiological aspects. Veterinary Research 34, 47-56.
131. Van Zeveren A. (2012). Veeteelt, deel 2 Bijzondere veeteelt: Hoofdstuk 9: het rund, melkvee.
Cursus Faculteit Diergeneeskunde, Gent, p14-15.
132. Veenhuizen J.J., Drackley J.K., Richard M.J., Sanderson, T.P., Miller L.D., Young J.W. (1991).
Metabolic changes in blood and liver during development and early treatment of experimental fatty
liver and ketosis in cows. Journal of Dairy Science 74(12), 4238-4253.
133. Ververs C., Hostens M., Caluwaerts T., de Kruif A., Opsomer G. (2010). Is er een verband tussen
het verloop van de aanvangsfase van de lactatiecurve en het optreden van de eerste oestrus post
partum bij hoogproductieve melkkoeien? Vlaams Diergeneeskundig Tijdschrift 79, 381-388.
134. Wada Y., Muto M., Matsuura K. (1995). Prognosis of cows with displaced abomasums and fatty
infiltration of the liver. Journal of the Japan Veterinary Medical Association 48, 387-390 (abstract in
Englisch).
135. Wildman E.E., Jones G.M. en Wagner P.E. (1982). A dairy cow body condition scoring system
and its relationship to selected production characteristics. Journal Dairy Science 65, 495-501.
136. Young J.WW. (1976). Gluconeogenesis in cattle: significance and methodology. Journal of Dairy
Science 60, 1-15.
137. Zhao F., Dixon W. T., Kennelly J.J. (1996). Localization and gene expression of glucose
transporters in bovine mammary gland. Comprehensive Biochemical Physiology 115, 127-134.
138. Zhao, F.Q., en Keating A. F. (2007). Expression and regulation of glucose transporters in the
bovine mammary gland. Journal of Dairy Science 90, 76-86.
139. Zulu V.C., Sawamukai Y., Nakada K., Kida K., Moriyoshi M. (2002). Relationship among insulin-
like growth factor-I, blood metabolites and postpartum ovarian function in dairy cows. Journal of
Veterinary Medicine Science 64, 879-885.



















