
Ctn Gn Exprn n th t d nd Spln f A/Al M Inftd th M lprila.ilsl.br/pdfs/v65n1a10.pdf · l 5 br I rntd...
10
Volume 65, Number I Printed in the U.S.A. (ISSN 0I48-916X) iNTERNAI IONAI. JOURNAI ()F LITROSY Cytokine Gene Expression in the Foot Pad and Spleen of BALB/cAJcl Mice Infected with M. leprae' Yasuko Yogi, Hiroko Nomaguchi, Yoshiki Sakamoto, Masanori Matsuoka, Takao Fujimura, Haruki Okamura, Kyoji Hioki, Muneo Saito and Tatsuji Nomura? The mechanisms of pathogenesis for Mycobacterium leprac infection remains unknown. Mice with cellular immuno- deficiency are highly susceptible to M. leprac which is an intracellular pathogen ( 4. 6. 13, 16, 22, 33, 49, 50 , ) In immunocompetent mice, the only susceptible sites to M. leprae infection are skin tissues, such as the foot pad and the ear. When M. leprae are inocu- lated into the foot pads of normal mice, the bacilli can multiply in the site hut the infec- tion is limited to 10' bacilli ( 32 ' 37-3"). Even though M. leprae were inoculated into nor- mal mice by intravenous injection, they were not able to multiply in the viscera such as the liver and the spleen ( 33 ). To elucidate the reasons for the restricted growth of M. leprae in the foot pads of immunocbmpe- tent mice, we examined and compared the expression of cytokine mRNAs in the foot pads and spleens of BALB/cAJcl mice in- fected with M. leprae. MATERIALS AND METHODS Mice. BALB/cAJcl mice were bred in the Central Institute for Experimental Ani- mals, Kanagawa, Japan. Twenty female mice aged 6 weeks were used in the experi- ' Received for publication on 18 September 1995; accepted for publication in revised form on 7 August 1996. = Y. Yogi, Ph.D.; H. Nomaguchi, Ph.D.; Y. Saka- moto, Ph.D.; M. Matsuoka, Ph.D., National Institute for Leprosy Research, 4-2-1 Aoba-cho, Higashimu- rayama-shi, Tokyo 189, Japan. T. Fujimura, Ph.D., De- partment of Dermatology, Kitasato University, School of Medicine, 1-15-1 Kitasato, Sagamihara-shi, Kana- gawa 228, Japan. H. Okamura, Ph.D., Department of Bacteriology, Hyogo College of Medicine, Nishi- nomiya, Hyogo 663, Japan. K. Hioki, 13.S.; M. Saito, B.S.; T. Nomura, M.D., Central Institute for Experi- mental Animals, 1430 Nogawa, Miyamite-ku, Kawa- saki-shi, Kanagawa 216, Japan. 80 ment. They were housed in the Animal Care Facility of our laboratory after inocu- lation with M. leprae. M. leprae. Leprosy bacilli, Thai-53 strain, derived from foot pad passage of nude mice were used. The suspensions of viable ( 3 ') or heat-killed M. leprae ( , D) were prepared as described by references indi- cated. Inoculation. Mice were infected with viable or heat-killed M. leprac each in both hind foot pads (2 x 10 7 ) combined with in- travenous inoculation at a dose of 4 x 10 7 . Two mice each were sacrificed on days 1, 10, 30 and 150 of inoculation, and the re- sults were compared with those of the mice prior to infection (24 hr before infection; day 0). Cytokine genes detection by RT-PCR. The expression of cytokine genes in the foot pads and spleens was examined by the reverse transcriptase-polymerase chain re- action (RT-PCR) method. RT-PCR of cy- tokine mRNAs from mouse tissues uninoc- ulated (day 0) or inoculated with M. leprae at varying periods of time as described above was performed as described ( 47 . 4 H). In brief, total RNA was extracted by the acid guanidinium method from 2 foot pads and 2 spleens which were isolated from 2 infected mice, pooled and frozen, and cDNA was synthesized using Moloney murine leuke- mia virus (M-MLV) reverse transcriptase and oligo (dT). Reactions were incubated in a thermal cycler (Astec High Voltage, Ash- land, Massachusetts, U.S.A.; PC-800) for 35 cycles. The cDNA concentrations were normalized to yield equivalent /3-Actin PCR products. After PCR, 10 pl of the DNA from each tube were loaded onto 1.5% agarose gels in TAE buffer. Products were visualized by ethidium bromide stain- ing. The sense and antisense printers used were as follows: Interleukin-1 alpha (IL-1-
Transcript of Ctn Gn Exprn n th t d nd Spln f A/Al M Inftd th M lprila.ilsl.br/pdfs/v65n1a10.pdf · l 5 br I rntd...

Volume 65, Number IPrinted in the U.S.A.
(ISSN 0I48-916X)
iNTERNAI IONAI. JOURNAI ()F LITROSY
Cytokine Gene Expression in the Foot Pad and Spleenof BALB/cAJcl Mice Infected with M. leprae'
Yasuko Yogi, Hiroko Nomaguchi, Yoshiki Sakamoto, Masanori Matsuoka, TakaoFujimura, Haruki Okamura, Kyoji Hioki, Muneo Saito and Tatsuji Nomura?
The mechanisms of pathogenesis forMycobacterium leprac infection remainsunknown. Mice with cellular immuno-deficiency are highly susceptible to M.leprac which is an intracellular pathogen( 4. 6. 13, 16, 22, 33, 49, 50 ,) In immunocompetentmice, the only susceptible sites to M. lepraeinfection are skin tissues, such as the footpad and the ear. When M. leprae are inocu-lated into the foot pads of normal mice, thebacilli can multiply in the site hut the infec-tion is limited to 10' bacilli ( 32 ' 37-3"). Eventhough M. leprae were inoculated into nor-mal mice by intravenous injection, theywere not able to multiply in the viscera suchas the liver and the spleen ( 33 ). To elucidatethe reasons for the restricted growth of M.leprae in the foot pads of immunocbmpe-tent mice, we examined and compared theexpression of cytokine mRNAs in the footpads and spleens of BALB/cAJcl mice in-fected with M. leprae.
MATERIALS AND METHODSMice. BALB/cAJcl mice were bred in
the Central Institute for Experimental Ani-mals, Kanagawa, Japan. Twenty femalemice aged 6 weeks were used in the experi-
' Received for publication on 18 September 1995;accepted for publication in revised form on 7 August1996.
= Y. Yogi, Ph.D.; H. Nomaguchi, Ph.D.; Y. Saka-moto, Ph.D.; M. Matsuoka, Ph.D., National Institutefor Leprosy Research, 4-2-1 Aoba-cho, Higashimu-rayama-shi, Tokyo 189, Japan. T. Fujimura, Ph.D., De-partment of Dermatology, Kitasato University, Schoolof Medicine, 1-15-1 Kitasato, Sagamihara-shi, Kana-gawa 228, Japan. H. Okamura, Ph.D., Department ofBacteriology, Hyogo College of Medicine, Nishi-nomiya, Hyogo 663, Japan. K. Hioki, 13.S.; M. Saito,B.S.; T. Nomura, M.D., Central Institute for Experi-mental Animals, 1430 Nogawa, Miyamite-ku, Kawa-saki-shi, Kanagawa 216, Japan.
80
ment. They were housed in the AnimalCare Facility of our laboratory after inocu-lation with M. leprae.
M. leprae. Leprosy bacilli, Thai-53strain, derived from foot pad passage ofnude mice were used. The suspensions ofviable ( 3 ') or heat-killed M. leprae ( ,D) wereprepared as described by references indi-cated.
Inoculation. Mice were infected withviable or heat-killed M. leprac each in bothhind foot pads (2 x 10 7 ) combined with in-travenous inoculation at a dose of 4 x 10 7 .Two mice each were sacrificed on days 1,10, 30 and 150 of inoculation, and the re-sults were compared with those of the miceprior to infection (24 hr before infection;day 0).
Cytokine genes detection by RT-PCR.The expression of cytokine genes in thefoot pads and spleens was examined by thereverse transcriptase-polymerase chain re-action (RT-PCR) method. RT-PCR of cy-tokine mRNAs from mouse tissues uninoc-ulated (day 0) or inoculated with M. lepraeat varying periods of time as describedabove was performed as described ( 47 . 4H). Inbrief, total RNA was extracted by the acidguanidinium method from 2 foot pads and 2spleens which were isolated from 2 infectedmice, pooled and frozen, and cDNA wassynthesized using Moloney murine leuke-mia virus (M-MLV) reverse transcriptaseand oligo (dT). Reactions were incubated ina thermal cycler (Astec High Voltage, Ash-land, Massachusetts, U.S.A.; PC-800) for35 cycles. The cDNA concentrations werenormalized to yield equivalent /3-ActinPCR products. After PCR, 10 pl of theDNA from each tube were loaded onto1.5% agarose gels in TAE buffer. Productswere visualized by ethidium bromide stain-ing. The sense and antisense printers usedwere as follows: Interleukin-1 alpha (IL-1-

65, 1^Yogi, et al.: Cytokine Gene Expression^ 81
a), 5 '-CTCTAGAGCACCATGCTACAG-AC-3' and 5'-TGGAATCCAGGGGAAA-CACTG-3'. IL-2, 5'-ATGTACAGCATG-CAGCTCGCATC-3' and 5 '-GGCTTGTT-GAGATGATGCTTTGACA-3'. IL-4, 5 '-A-TGGGTCTCAACCCCCAGCTAGT-3 ' and5 '-GCTCTTTAGGCTTTCCAGGAAGTC-3 '. IL-6, 5 '-ATGAAGTTCCTCTCTGCA-AGAGACT-3' and 5 '-CACTAGGTTTGC-CGAGTAGATCTC-3 '. IL-10, 5 '-TACC-TGGTAGAAGTGATGCC-3' and 5'-CA-TC ATGTATGCTTCTATGC-3 '. IL-12 (p40),5 '-CAGAAGCTAACCATCTGGTTTG-3 'and 5 '-TCCGGAGTAATTTGGTGCTTC-ACAC-3'. Gamma interferon (IFN-y), 5'-TGAACGCTACACACTGCA TCTTGG-3 ' and 5'-CGACTCCTTTTCCGCTTCCT-GAG-3 '. Tumor necrosis factor-alpha(TNF-a), 5 '-GGCAGGTCTACTTTGG-AGTCATTGC-3' and 5'-ACATTCGAG-GCTCCAGTGAATTCGG-3'. TNT-A 5'-TGGCTGGGAACAGGGGAAGGTTGAC-3' and 5'-CGTGCTTT CTTCTAG AAC-CCCTTGG-3'. CD4, 5 '-TGTGCCGAGC-CATCTCTCTTAGG-3 ' and 5 '-GCACTG-AGAGTGTCATGCCGAAC-3'. CD8, 5'-ATGCAGCCATGGCTCTGGCTGG-3 'and 5'-GCATGTCAGGCCCTTCTGGG-TC-3'. /3-Actin, 5 '-TGGAATCCTGTGG-CATCCATGAAAC-3' and 5 '-TAAAACG-CAGCTCAGTAACAGTCCG-3'.
Immunohistologic staining of mousefoot pad tissues. Lymphocytes bearingCD4 and CD8 surface expression in thefoot pad prior to the inoculation and 30days after viable M. leprae inoculationwere determined by immunohistochem-istry. The tissue with OCT medium (Miles,Inc., Elkhart, Indiana, U.S.A.) was cut for6-pm thickness, fixed with acetone andchloroform, and then blocked with normalrabbit serum (Dako Corp., Carpenteria,California, U.S.A.; X902) before undergo-ing incubations with the monoclonal anti-bodies (anti-CD4 and anti-CD8; Serotec,Kidlington, U.K.; KT174 and KT15)overnight at 4°C followed by biotinylatedrabbit anti-rat immunoglobulins (DakoE0468) for 30 min. Slides were washedwith phosphate buffered saline (PBS) be-tween incubations. Primary antibodies werevisualized by using the LSAB kit (Dako).Slides were counterstained with hema-toxylin and mounted in Ukitt (0. Kinder,Germany).
ELISA. The mouse serum immunoglob-ulin of each group was measured by anELISA method using goat anti-mouse IgGand IgM. The serum antibodies to M. lep-rae phenolic glycolipid-I (PGL-I), lipoara-binomannan-B (LAM-B) and heat-shockprotein 65-kDa (hsp65 kDa) were measuredby the ELISA method of Cho, et al. ( 5 ).LAM-B was prepared in our laboratoryfrom a M. leprae cell-wall fraction obtainedfrom armadillo tissues ("); PGL-I also wasprepared from M. leprae-infected armadillotissues using the method by Hunter, et al.co. I 2 ,
) M. leprae-hsp65 was also preparedin our laboratory from an Escherichia colistrain and affinity-purified with monoclonalantibody to hsp65 ( 2"). Mouse serum wasdiluted at 1:100 with 2% bovine serum al-bumin-phosphate buffered saline-azide(BSA-PSA-azide) for antibody titration.The microplates (Nunc, Roskilde, Den-mark) were coated with 50 pl/well of PGL-I at 2 pg/ml, LAM-B at 1 pg/ml or hsp65 at2 pg1m1 in 0.05 ml carbonate buffer (pH9.6); the other half of each plate was coatedwith a coating buffer alone as a control. Theperoxidase-conjugated goat anti-mouse IgGF(ab ')2 and IgM F(ab ')2 fragment (diluted1:1000) was used for secondary antibody.Serum antibody level (01) 4 ,,„_ 6,„„„,) was cal-culated by subtracting the mean opticaldensity (OD) of the control wells from themean OD of the antigen-coated wells.
RESULTSCytokine mRNA expression in foot
pads of M. /eprae-inoculated mice. Thechanges in the cytokine mRNAs expressionover time in the foot pads of uninoculated(24 hr before infection; day 0) and intra-venously inoculated mice combined withthe foot pad injection with viable or heat-killed M. leprae are shown in Figure 1.Physiologic expression of IL-la andTNF-13 mRNAs were observed in the footpads of the uninoculated mice. In addition,in mice inoculated with viable M. leprae,the usual appearance of IL-la and TNF-PmRNA was intensified; TNF-a mRNA wasinduced on day 1 of inoculation. The ex-pression of these mRNAs increased withthe progression of the infection until day10, and then decreased on day 30 and in-creased again on day 150. The same patternof mRNA expression was also observed in

82^ International Journal of Leprosy^ 1997
1 2 3 4 5^6 7 8 9C M^1 2 3 4 5^6789 M
A
B
C
E
F
G
H
FIG. I. Cytokinc niRNA expression in foot pads ofAl. /cm -tie-inoculated mice. PCR analysis of cDNAfrom two foot pad tissues pooled from 2 infected micewas carried out at varying times by: lane I, 24 hr be-fore infection (day 0); lane 2, on day 1; lane 3. on day10; lane 4, on day 30: lane 5, on day 150 after infectionwith viable Al. leprae. Lane 6, on day I; lane 7. on day10; lane 8, on day 30; lane 9, on day 150 after infectionwith heat-killed Al. leprae. Lane C was the positivecontrol from ConA-stimulated spleen cells. Specificprimers used for the sequence of cytokines, CD4 andCD8 were : A = /3-Actin, 13 = CD4, C = CDS, D = IL-
E = IL-2, F = G = TNF-a and II = TNF-13.Reactions were incubated in an Astec-800 for 35 cy-cles as conditions with denaturation I min, 94°C; an-nealing 2 min, 60°C or 65°C: extension 3 min, 72°C.
mice inoculated with heat-killed M. l('prae;the degree of induction of these mIZNAswas lower on day I and the expression wasslightly stronger after day It).
A ••1.•• •.••
B^V if^111 -
C^11"/
D^um. ere^up le
wir^-^ el•
Iry
H -^1p
I^111/ -^- 11. isr e
up IMP I.^v
K^- gip
Wipe/ OP
FIG. 2. Cytokine niRNA expression in the spleensof Al. hp/we-inoculated !nice. PCR analysis of cDNAfirom 2 spleen tissues pooled from 2 infected mice werecarried out at varying times as well as the foot pad afterinoculation with viable or heat-killed Al. leprae. Lane I ,24 hr before inoculation (day 01; lane 2, on day I; lane1 on day 10: lane 4, on day 30; lane 5, on day 150 afterinoculation with viable Al. hprae. Lane 6, on day I;lane 7, on day 10: lane 8, on day 30: lane 9, on day 150after inoculation with heat-killed M. /eproe. Specificprinters used were as follows: A = /3-Actin. 13 = CD4, C= CD8, D = = IL-2, F=11,-4, G = 1L-6. 11 =
= IL-12(p40). J =^K = TNF-a, L = TNF-0.

65, I^Yogi, et al.: Cvtokine Gene Expression^ 83
Cylokine mRNA expression in spleensof M. /cpme-inoculated mice. The changesof cytokine inRNA expression in the spleenover time in uninoculated mice (day 0) andmice inoculated with viable or heat-killedM. leprae at varying periods of 1, 10, 30and 150 days are shown in Figure 2. All ofthe cytokine inRNAs were examined; IL-1 a, -2, -4, -6, -10, -12, 1FN-y, TNF-a andTNF-f3 were expressed physiologically inthe spleens of uninoculated mice 24 hrbefore infection. CD4 and CD8 mRNAsalso were expressed physiologically. Inmice inoculated with viable M. /cynic, CD4inRNAs were undetectable on day I of in-oculation, but the expression was recoveredover time. In mice inoculated with heat-killed M. leprae, CD4 mRNA expression inthe spleen was decreased up to day 30 of in-oculation, and 1L-2 and IL-6 inRNAs (ex-cept for a weak expression on day 1) werenot expressed. An interesting observationwas that the loss of CD4 mRNA expressionon day I of inoculation of live M. lepraewas accompanied by a diminished expres-sion of IL-2, -4, -6 and IL-12 mRNAs, andthe fluctuation in cytokine mRNA expres-sion followed the pattern of fluctuation ofCD4 mRNA expression. An increase in IL-4 and IL-10 mRNAs expression was ob-served during the course of infection inmice inoculated with viable M. leprae.Fluctuation of IL - 1 - a mRNA expressionwas observed (luring the course of infectionin mice inoculated with live bacteria but notin those inoculated with killed bacteria.TNF-/3 mRNA expression remained con-stant at the pre-inoculation level in bothgroups inoculated with viable and heat-killed M. lepme.
Immunohistological examinations OfC1)4+ and CD8+ lymphocytes in footpads of M. kprac-infected mice. On day30 of inoculation the foot pads received vi-able M. leprae and the foot pads prior to theinoculation were stained immunohistologi-cally to examine for T-cell induction due toM. leprae infection. CD4+ and CD8+ lym-phocytes were detected in the subcutaneoustissue and in the intermuscular layer asshown in Figure 3 but were not observedprior to the inoculation of M. leprae (datanot shown).
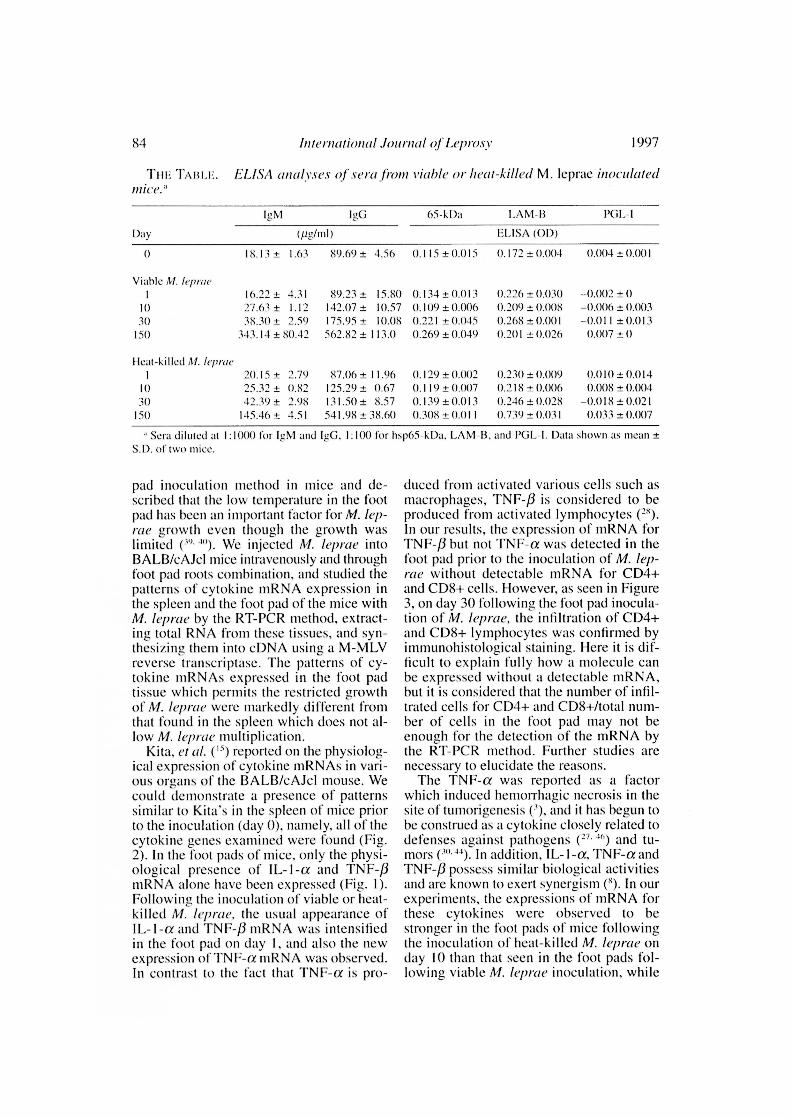
LISA analyses of sera from M.leprae-inoculated mice. Increases in the
FIG. 3. Immunohistological stains of (71)-1 andCD8 lymphocytes in foot pads of a mouse on day 30 ofinfection with viahle M. leprae showing CD4+ (A) andCDS+ (B) lymphocyte infiltration into the intermuscu-lar layer (x400).
IgM and IgG levels in the serum, due pre-sumably to M. /epray inoculation, were ob-served 10 days after inoculation of M. lep -
Itle (The Table). There were no differencesin increase of the antibody response to re-combinant hsp65 between the groups in-fected with viable and heat-killed M. lep-rae. The antibody level to LAM-I3 antigenwas evidently higher in the group inocu-lated with heat-killed M. /cpme after 150days of inoculation. No significant in-creases in antibody responses to PGL - Iwere observed in either group infected withviable or killed bacteria after 1St) days ofinoculation.
DISCUSSIONSince Hansen first described M. leprae as
the pathogenic organism of leprosy, numer-ous animal experiments have been con-ducted ( 21 . 21. Shepard developed the foot
84^ International Journal of Leprosy^ 1997
Tin: TABLE. EL/SA ana/yses of sera from viable or heat-killed M. leprae inoculatedmice."
Day
IgNil IgG 65-k1)a LAM-13 PGL-1
(t1g/m1) [LISA (0D)
0 18.13±^1.63 89.69 ±^4.56 0.115±0.0150.015 0.172 ± 0.004 0.004 ± 0.001
Viable M. hproe
1 16.22 ±^4.31 89.23 ±^15.80 0.134 ± 0.013 0.226 ± 0.030 -0.002 ± 010 27.63 ±^1.12 142.07 ±^10.57 0.109 ± 0.006 0.209 ± 0.008 -0.006 ± 0.00330 38.30 ±^2.59 175.95 ±^10.08 0.221 ± 0.045 0.268 ± 0.001 -0.011 ± 0.013
ISO 343.14 ± 80.42 562.82 ± 113.0 0.269 ± 0.049 0.201 ± 0.026 0.007 ± 0
1 teat-k il led M. frprac
1 20.15 ±^2.79 87.06 ± 11.96 0.129 ± 0.002 0.230 ± 0.009 0.010 ± 0.01410 25.32 ±^0.82 12y9± 0.67 0.119 ± 0.007 0.218 ± 0.006 0.O08±0.0040.00430 42.39 ±^2.98 131.50 ±^8.57 0.139 ± 0.013 0.246 ± 0.028 -0.018 ± 0.021
150 145.46 ±^4.51 541.98 ± 38.60 0.308 ± 0.011 0.739 ± 0.031 0.033 ± 0.007
Sera diluted at 1:1000 for IgM and leG, 1:100 for hsp65-kDa, LAM-IL and PGL-1. Data shown as mean ±S.D. of two 'nice.
pad inoculation method in mice and de-scribed that the low temperature in thepad has been an important factor for M. lep-rae growth even though the growth waslimited ("' 5 )). We injected M. /eprae intoBALB/cAJcl mice intravenously and throughfoot pad roots combination, and studied thepatterns of cytokine mRNA expression inthe spleen and the foot pad of the mice withM. leprae by the RT-PCR method, extract-ing total RNA from these tissues, and syn-thesizing them into cDNA using a M-MLVreverse transcriptase. The patterns of cy-tokine mRNAs expressed in the foot padtissue which permits the restricted growthof M. leprac were markedly different fromthat found in the spleen which does not al-low M. /eprae multiplication.
Kita, et al. ( 15 ) reported on the physiolog-ical expression of cytokine mRNAs in vari-ous organs of the RALB/cAJcl mouse. Wecould demonstrate a presence of patternssimilar to Kita's in the spleen of mice priorto the inoculation (day 0), namely, all of thecytokine genes examined were found (Fig.2). In the foot pads of mice, only the physi-ological presence of IL-1-a and TNF-/3mRNA alone have been expressed (Fig. 1).Following the inoculation of viable or heat-killed M. /eprae, the usual appearance ofIL-1-a and TNF-f3 mRNA was intensifiedin the foot pad on day 1, and also the newexpression of TNF-a tuRNA was observed.In contrast to the fact that TNF-a is pro-
duced from activated various cells such asmacrophages, TNF-P is considered to beproduced from activated lymphocytes (").In our results, the expression of mRNA forTNF-/3 but not TNF-a was detected in thefoot pad prior to the inoculation of M. lep-rae without detectable mRNA for CD4+and CD8+ cells. However, as seen in Figure3, on day 30 following the foot pad inocula-tion of M. leprae, the infiltration of CD4+and CD8+ lymphocytes was confirmed byimniunohistological staining. Here it is dif-ficult to explain fully how a molecule canbe expressed without a detectable mRNA,but it is considered that the number of infil-trated cells for CD4+ and CD8+/total num-ber of cells in the foot pad may not beenough for the detection of the mRNA bythe RT-PCR method. Further studies arenecessary to elucidate the reasons.
The TNF-a was reported as a factorwhich induced hemorrhagic necrosis in thesite of tumorigenesis ( 1 ), and it has begun tobe construed as a cytokine closely related todefenses against pathogens ( 27-') and tu-mors 0 44). In addition, IL-1-a, TNF-a andTNF-P possess similar biological activitiesand are known to exert synergism ( 5 ). In ourexperiments, the expressions of mRNA forthese cytokines were observed to bestronger in the foot pads of mice followingthe inoculation of heat-killed M. leprac onday 10 than that seen in the foot pads fol-lowing viable M. /eprae inoculation, while

65, I^Yogi, et al.: Crtokine Gene Expression^ 85
the appearance of these cytokines was notchanged in the spleens of mice inoculatedwith either viable or heat-killed M. leprae(Figs. 1 and 2).
The changes in the serum antibody titersof infected BAL13/cAJcl mice were foundfrom day 10 following the infection of M.leprae for the amount of IgM and IgG, andboth of the specific antibodies againstlisp65 kDa and LAM-13 were found to in-crease when compared with titers prior tothe infection. The titers of LAM-13 werehigher in the mice inoculated with heat-killed M. /eprae than in those receiving vi-able bacteria on day ISO as shown in TheTable. Although the elevation of the anti-body level against PGL-I was not observed,the presence of PG L-1 was confirmed byinimunohistological stain in the tissues ofviable M. /eprae infection (data not shown).PGL-I ( 1 '. 2 `. 25 . 5 ) and LAM-13 (H-' 2 ) are re-ported to be immunosuppressive antigensfor cellular immunity, and hsp65 ( 2 $) is re-ported to be a highly immunogenic antigenfor Immoral and cellular immunity. Whenthe foot pads of the BA1.13/cAJcl (en-thymic) mice were inoculated with a largeamount of M. leprae, the bacilli remain I'ora period of time in the inoculation site ofthe foot pad. demonstrating the establish-ment of persistent and resistant infection,even though there is no increase in thenumber of bacilli as reported by Rees (In the foot pad, cytokine production consid-ered to eliminate the leprosy bacilli may besuppressed 1G PGL-I which is producingand releasing into tissues by viable M. lep-rae. In fact, PGL-I was detectable in thefoot pad inoculated with M. leprae by im-munohistological examination as men-tioned above, and the antibody levels toLAM-B were evidently higher in the serumof the infected mice than those prior to theinoculation.
In contrast to the foot pad, all of the cy-tokine genes tested appeared in the spleen,except the disappearance of CD4 mRNA onday I of M. leprae infection accompanyingthe reduced expression of IL-2, -4, -6, andIL-12 mRNAs. The CD4 mRNA expres-sion was recovered on day 10 of infectionwith viable M. /eprae, with a correspondingincrease in lymphokine inRNA expressionwhich had been decreased on day 1. Inother words, following the infection with
M. leprae, the expression of CD4 mRNA islost and reduced immediately: at the sametime the reduction of the appearance oflymphokine genes occurs.
The results of the present study suggestthat M. leprae exert a direct effect on CD4lymphocytes which is related to inhibitingthe expression of cytokine genes for theelimination of bacilli from the host tem-porarily. The temporary loss of expressionof CD4 niRNA and the loss of accompany-ing cyktokine mRNA expressions in thespleen were stronger by the killed M. lepraeinoculation than by the viable bacilli in thespleen. In the case of the killed bacilli inoc-ulation, they never recovered up to 30 daysof the post-inoculation, and the expressionfor IL-2 and 11,-6 mRNAs never recovered,even the end of this experiment (Fig. 2).Based on these facts for cytokine patterns,in addition to the low temperature of thefoot pad, as described by Shepard (''''.")), itis considered that a long-term continuationspecific to M. leprae infection is establishedin the foot pad.
Mitsuyama, et al. explained that killedbacteria have a weak ability to induce IL-Iproduction in macrophages so that T-cellactivation cannot proceed, leading to a fail-ure in eliciting delayed-type hypersensitiv-ity (DTI-I) In M. leprae infection, how-ever, IL-1-a niRNA was detected in boththe foot pads and the spleens of mice inoc-ulated with killed M. /eprae the same as vi-able M. /eprae in our results shown in Fig-ures I and 2. On day 10 after infection, theexpression of IL-1-a mnRNA was strongerin the foot pad by the killed M. /gave in-fection. It has been reported that the DTI-Ireactions have been induced in mice pre-in-oculated with killed ( 17 ) or viable M. leprae( 2 ") by challenge with M. leprae lysate.
Recently, the analysis for the experimen-tal results using TNF-a knockout mice clar-ified the role of this cytokine against Ultra-cellular parasites. When TNILa p55 knock-out mice were inoculated intravenouslywith Lvsteria moriorytogenes, all of theknockout mice died within 6 days after theinoculation while all of the normal controlmice were alive (H However, theseknockout mice were normal against virusinfection, showing normal function for cy-totoxic lymphocytes. There is a reportthat the administration of 11,-1-a into mice

86^ Thternationa/ Journal al Leprosy^ 1997
inhibit the growth of intracellular parasites('). In the intravenous inoculation of Lyste-ria, 90% of them are distributed in the liver,then killed and removed by Kupffer's cells.Thus, recent studies for the understandingof cellular parasitic infections indicated thatnonspecific immunoregulation of macro-phages is more important than the antigen-specific immune system by T lymphocytes
` 5 ). In the case of M. leprae which cannotmultiply in the visceral organs, such as thespleen, similar removal mechanisms may alsobe established by the estimation of TNILamRNA patterns in our results (Figs. I and 2).
The cytokine genes, IL-1-a, TNF-a andTNF-/3 mRNAs, which express in the footpad of immunocompetent BALI3/cAJclmice infected with M. leprae were also ob-served in the foot pad of SCID and nudemice which are highly susceptible to M.leprae ( 5 '). It has been reported that IL- Ihas the ability to promote the growth ofpathogenic Escherichia colt ("). Beutler, eta!. (') have reported that a high degree ofhomology exists between the TNF andmacrophage-secreted factor cachectin, andKawakami and Ilayata (") have reportedthat TNF may be a potent cachexia or au-toimmune inducing agent for the 'willfulexcess of TNF production and, also, Amiri,et al. (') have reported that schistosome uti-lizes TNF to maintain its progeny afteroviposition. TNF-/3 also has been shown tobe an important factor in the construction oflymph nodes (."). In the mechanisms of M.leprae infection and immunity, whetherthese cytokines play only a role in the elim-ination of M. leprue remains to be eluci-dated. In immunocompetent mice, the onlysites that are susceptible to M. leprae infec-tion are skin tissues, including the foot pad.Although the significance of the expressionof IL-1-a, TNF-a and TNF-/3 mRNAs inthe foot pads of these mice cannot be ascer-tained without further studies, our presentstudy suggests that tissue-specific, local, im-munologic characteristics are important inM. leprae growth, and the overall immuno-logic competence of the infected host exertsfurther influence on its multiplication.
SUMMARY
The cytokine mRNAs expressed in thefoot pads and spleens of BALB/cAJcl mice
infected with Mycobacterium leprue werestudied by the reverse transcriptase-poly-merase chain reaction (RT-PCR) methodusing cytokine-specific primers for inter-leukin-1 alpha (IL-1a), -2, -4, -6, -10, -12-(p40), gamma interferon (IFN-y), tumornecrosis factor-alpha (TNF-a), and TNILP,and then for CD4 and CD8 markers. Thepattern of cytokine gene expression in thefoot pad which supports M. leprue growthwas different front the expression in thespleen which does not permit M. lepraemultiplication in mice. 13efore BALB/cMclmice were infected with M. leprac, IL-laand TNF-/3 mRNAs were expressed physi-ologically in the foot pad while all of thecytokine genes examined were expressed inthe spleen. In the foot pads of mice inocu-lated with M. leprae, in addition to the phys-iological appearance of IL- I a and TNF-/3mRNAs, these signals were intensified.TNF-a expression was induced by the in-fection. On the other hand, in the spleens ofmice inoculated with M. leprac, CD4mRNA expression disappeared on day I ofthe infection, which was accompanied by thereduced expression of IL-2, -4, -6, and -12mRNAs. The recovery of CD4 mRNA ex-pression at a latter stage was accompaniedby a corresponding increase of the cytokinemRNA expression. It was suspected thatthese results might permit restricted growthof M. leprae in the foot pads of normalmice. Furthermore, our study suggests thattissue-specific, local, immunologic charac-teristics are important in M. leprae growth.
RESUNIEN
utilizando to reaccion en cadena de la polimerasa-transcriptasa reversa (12T-PCR) se estudiaron losmRNAs para citocinas expresados en as almohadillasplantares de ratones It ALB/cMcl infectados con My-cobacterium leprae. Se utilizaron sondas (iniciadores)para interteucina-t alfa (IL-M, IL-2, IL-4, IL-6, IL-10,IL-12 (p4(), interferón gamma (IFIN-)), factor denecrosis tumoral alfa (TNIFtx), TNF/3, y para los mar-cadores CD4 y CD8. El pzitrán de expresion de losgenes para citocinas en la ahnohadilla plantar (que per-mite el crecimiento de Al. leprae) file di ferente del pa-trim de expresión de citocinas en el bait) (el cual nopermite la multiplicación de Al. leprac en el ratón).Mientras que mil() los mRNAs para IL-1 a y TNF/3 seexpresaron tisiológicamente en las almohadillasplantares de los ratones 13AL13/cAJC1 antes de su in-fecchin con Al. leprac, todos los genes para citocinasexaminados fueron expresados en los hams. En as

65. I^Yogi, et al.: Cvtokine Gene Evnvssion^ 87
almohadil las plantares de los ratones inoculados conAl. /eprae, adeintis de la •parichin lisioltigica (le los
niRNAs para IL- I a y TN113, estas seriales rucron in-
tensilicadas.1,a expreshin (le TNEa foe inducida por la
Por otro Lido, en los hares (le los animalesinoculados con M. teprac. la expreshin del inIZNApara CO4 desaparcei() hacia el dia I (1c 1.1 infecchin y
esto sC acomparui (le la expreshin reducida (le los inR-
NAs para 11,-2,11.-4,11.-6, c 11,-12. La recuperachin (lela expreshin del niIZNA para CD4 en los estadios Inas
avail/tidos (le la inlecchin, SC 1.1COillpallti del correspon-
theme increment() en la expresitin (le los tiWNAs paralas otra.s citoeinas. Estos resultados podrían estar rela-ciuna(Ios con el crecimiento restringido de M. /eprae
en Its almoltadillas plantares (le los mimics normales.
Adeintis, !mew() estudio sugicre (Fie las caracteristicaslocales y especilicas de tejido influyen importante-mente en el crecimiento de ,1/. teprae.
Les A 1:Nni de cytokine exprinit:s dims lesc(pssinets plantaires et les rates de souris BALII/cAJclinfectes par Mycobacterium teprae out etc (!tudit::s parla ini:thode de reaction reverse de transcripase-poly-merase en Chaise (RT-P('R), en utilisant des mar-queurs spt:cifiques de c) tokines pour l'interleukinc-1
alpha (11.-1a), -2, -4, -6, -10, - I 2(p-10), gamma inter-
ft:ron (IEN-y), le facteur de ni2crose tumorale alpha
(ENT-a), et le FNT-P, et pour les inarqueurs C1)4 et
CI)8. Le mode d'expression du gene (le cytokine dans
le coussinct plantaire qui supporte la croissance de M.//Trite aait different (le l'expression dans la rate qui nepermit pas la multiplication de .1/. /e/true clic/ lasouris. Avant (see les solids LBA:AJCI 11C SOielll 01-fecti.es par teprae. les AILNin (111.-la et ENT-0oat etc expriniCs physiologiquement dans le coussinet
plantaire alors que taus les g&nes de cytokines exam-nes etc exprints dans 1.1 rate. Dans les coussinetsplantaires (les souris inocult:ses :arc Al. leprae, enplus de l'apparition physiologique des Al(Nnt (111,-la
et ENT-/I, ces sign:tux ant etc intensiUs. (.'expressionde PNT-tx etait imitate par [infection. IYantre part,dans les raleS 5011115 infeeteCS 11L11 - ieprae, [ex-pression (le l'AIZNin de C1)4 disparut au premier jour(le l'infect ion, qui t:tait ticcompagnte dune reduction
(le [expression des AltNin (111.-2, -4, -6 et -12. Le
rétablissement de [expression de l'ARNin (le CI)4 ittin static plus tardif isms :tccompagml: (I' une augmenta-
tion correspondante (le [expression (le l'AIZNin (le cy-
tokine. On it soupeonnt: que ces resultats pourraientpermettre tine croissance IimiteC (le Al. leprae dons lescoussincts plantaires souris normales. 1)e plus.
noire antic suggere title les carack:ristiques locales,spt:cifiques par tissu, sow importantes pour 1.t crois-sauce (le Al. /t/,r/C.
AcknoNl ledgments. We thank Professor Robert I,.Modlin and his staff, at I.C.L.A., Los Angeles, C'ali-
fornia. I I.S.A., for RT-PC'R method. This work was
supported by a grant 116111 the Sasakawa MemorialIlealth Foundation.
REFERENCESI. ANIIR1, P., LocKsirx, R. NI, PARsi,ow, T. G.,
SAtucK, NI., REcrott, 12.144ER, D. and McKER-
tutw, J. II. Tumour necrosis factor a restores gran-
ulomas and induces parasite egg-laying in schisto-
some-infected SCID mice. Nature 356 (1992)604-607.
2. 13Eirri.m, 13., GREENwALD, D., HuLkuis, J. D.,C'ilAN0, M., PAN, Y.-C. E., NIATfitsoN, J., ULE-
vrrcit, R. and CD:Arof, A. Identity of tumour
necrosis factor and the macrophage-secreted Lie-tor cachectin. Nature 316 ( 1985) 552-554.
3. CARsvcD.I., h. A., OD), L. J., KAssEi., R. I., ET AL.An endotoxin-induced serum factor that causesnecrosis of tumors. Proc. Natl. Acad. Sci. U.S.A.72 (1975) 3666-3670.
4. Oulu., S., Rutty, J., Jon, C. K. and HASTINGS, R.C. The growth of :Mycobacterium /quite in nude
Lepr. Rev. 54 (1983) 283-304.
5. Clio, S.-N., YANAfillIARA, D. L., HUNTER, S. W.,
Gbirtyk, R. II. and BRENNAN, P. J. Serologicalspecificity of phenolic glycolipid- I from My-
cobacterium /tyro(' and use in serodiagnosis ofleprosy. Infect. Immun. 41 (1983) 1077-1083.
6. Col.sT08, M. J. AND 1111.SON, G. R. F. Growth of
M. teprac and M. marinum in congenitally
athymic (nude) mice. Nature 262 (1976) 399-401.7. Cir IITYNSKI, C. J. and litrowN, J. F.
111I1Fillc interleukin-1 a enhancement of nonspe-
cific antibacterial resistance. Infect. Inunun. 55
(1987) 2061-2065.8. DtivARD.1.0, C. A. Interleukin-1 and its biological
related cytokines. Adv. Immunol. 44 (1989)153-2(15.
9. DREvrrs, I). A., LEENEN, P. J. and CANIPBELL, P. A.Complement receptor type 3 (C1)11b/C1)18) in-volvement is essential for killing of Li.rieriamonorytogencs by Molise macrophages. J. 111)-1)11111ot. 151 (1993) 5-131-5439.
10. I luNnit, S. W. and BRENNAN, P. J. A novel phe-nolic glycolipid front Myeebacterhun hprae pos-
sibly involsed in immunogenicity and pathogenic-
ity. J. liacteriol. 147 ( 1981) 728-735.11.1Ium IA:, S. W., Giyitau), II. and BRENNAN, P. J.
Structure and tintigenicity of the phosphorylated
lipopolysaccharide antigens front the leprosy andtubercle bacilli. J. Biol. Chem. 261 ( 1986) 12345-12351.
12. I It irsirrit, S. W., STEwART, 13. S. and BRENNAN, P. J.Purification of phenolic glycolipid I from ar-madillo and 101111:10 sources. Int. J. Lepr. 53 (1985)484-487.
13. IsitAQui ,,, NI. Inoculation of loot pads of severe
combined immunodeficient mice with M. lep•ae
Int. J. Lepr. 60 (1992) 661.
1.1. K. \\\..0:.\ NIL NI. and 11AYATA, K. [UNE' Med. Ini-lnun01. 21) (1990) 615-620.

88^ International Journal of Leprosy^ 1997
21. NAKAMURA, NI. [Mycobacterium /civet(' and M.lepraemurium.] In: [Experimental Aninuils forLeprosy.] University of Tokai Press, Inc. (1985)Chapter 5, pp. 210-283.
22. NAKAMURA, K. and YoGi, Y. The nude mouse asan experimental lepromatous leprosy model: theenhancing effect of thymus cells in infected nudemice. (Abstract) Int. J. Lepr. Int. Stipp'. (1979)338.
23. NEILL, M. A. and KLEBANOFF, S. J. The effect ofphenolic clycolipid-1 from Mycobacterium lepraeon the antimicrobial activity of human macro-phages. J. Exp. Med. 167 (1988) 30-42.
24. Nomouctit, H. ]Heat shock protein and immu-nity.] Jpn. J. Lepr. 64 (1995) 188-199.
25. NoNiAcium, II., Dow, Y., OHNo, N., ET AL. Sup-pression of ConA responses of mouse lympho-cytes with unique M. leprae glycolipid. Jpn. J.Lepr. 58 (1989) 191-196.
26. NOMAGUCHI, H., MATSUOKA, M., KOHSAKA, K.,NAKATA, A. and huff, T. Overproduction, affinitypurification and characterization of 65-KDa pro-tein of Mycobacterhon le/we in Escherichia coll.Int. J. Lepr. 57 (1989) 817-824.
27. Otn, L. J. Tumor necrosis factor (TNF). Science230 (1985) 630-632.
28. PAUL, N. L. and RUDDLE, N. H. Lymphotoxin.Annual Rev. Immunol. 6 (1988) 407-438.
29. PETER, A. S. and JOHNSTONE, M. A. The search foranimal models of leprosy. Int. J. Lepr. 55 (1987)535-547.
30. PHILIP, R. and EPSTEIN, L. B. Tumour necrosisfactor as immunotnodulator and mediator of
monocyte cytotoxicity induced by itself, y-inter-feron and interleukin-1. Nature 323 (1986) 86-89.PEEEEER, K., MArstJYANtA, T., KUNDIG, T. M., ET
AI.. Mice deficient for the 55KD tumor necrosisfactor receptor are resistant to endotoxic shock,yet succumb to L. 0101101viogenes infection. Cell73 (1993) 457-467.Reis, R. J. W. Limited multiplication of acid-fastbacilli in the foot pads of [nice inoculated withMycobacterium hymn,. Br. J. Exp. Pathol 45(1964) 207-218.
33. REES, R. J. W., WATERS, M. F. R., WEDDELL, A. G.M. and PALMER, E. Experimental lepromatousleprosy. Nature 215 (1967) 599-602.
34. RoRAr, R., CLARK, B. I)., WoEFE, S. M. andDINARELLO, C. A. Enhancement of growth of vir-ulent strains of Escherichia co/i by interleukin-1.Science 254 (1991) 430-432.
35. ROSIN, H., GmDoN, S. and NORTH, R. J. Exacerba-tion of murine listeriosis by a monoclonal antibodyspecific for the type 3 complement receptor ofmyclo-monocytic cells. J. Exp. Med. 170 (1989)27-37.RoTHo, J., LESSLAUER, W., LOTSCHER, 11., ET AL.
Mice lacking the tumour necrosis factor receptor Iare resistant to TNF-mediated toxicity but highlysusceptible to infection by Listed(' monocyto-genes. Nature 364 (1993) 798-802.
37. SHPARD, C. C. Acid-fast bacilli in nasal excre-tion in leprosy, and results of inoculation of mice.Am. J. Hyg. 71 (1960) 147-157.
38. SHEPARD, C. C. The experimental disease that fol-lows the injection of human leprosy bacilli intofoot pads of mice. J. Exp. Med. 112 (1960)445-154.
39. SHEPARD, C. C. Multiplication of Mycobacteriumleprae in the foot pad of the mouse. Int. J. Lepr. 30(1962) 291-306.
40. SHEPARD, C. C. Considerations of the applicationof the foot pad technic in leprosy research. Int. J.Lepr. 33 (1965) 657-661.
41. SHILEY, L. D., ADAms, L. B. and KRAIIENBUHL, J.L. Inhibition of interferon-gamma-mediated acti-vation in mouse macrophages treated with lipoara-binomannan. Clin. Exp. Immunol. 80 (1990)141-148.
42. SIBLEY, L. D., HUNTER, S. W., BRENNAN, P. J. andKRAIIENBUHL, J. L. Mycobacterial lipoarabino-mannan inhibits gamma-interferon-mediated acti-vation of macrophages. Infect. Immun. 56 (1988)1232-1236.
43. Thom, P. D., GOELLNER, J., RuDDLE, N. H.,STREETER, P. R., FICK, A., MARIATHASAN, S.,
SMITH, S. C., CARISON, R., SHORNICK, L. P.,SCHOENBERGER, J. S., RUSSELL, J. H., KARR, R.and CHAPLIN, D. D. Abnormal development ofperipheral lymphoid organs in mice deficient inlymphotoxin. Science 264 (1994) 703-707.
44. URBAN, J. L., SHEPARD, II. M., ROTHSTEIN, J. L.and SUGARSIAN, B. J. Tumor necrosis factor: a po-tent effector molecule for tumor cell killing by ac-
15. KrrA, M., TONG, L. J., TANAKA, K. and ImANistti, J.Expression de l'ARN messager des cytokines
^
chez la souris clans des conditions physiologiques.^31.C. R. Soc. Biol. 187 (1993) 414-419.
16. KOIISAKA, K., Moto, T. and Iron, T. Lepromatoidlesion developed in nude mouse inoculated withM. leprae. Lepro 45 (1976) 177-187.
17. LAMB, F. 1., KINGSTON, E., ESTRADA-G, I. and^32.Cot.sToN, M. J. I leterologous expression of the65-kilodulton antigen of Mycobacterium lepraeand marine T-cell responses to the gene product.Infect. Immun. 56 (1988) 1237-1241.
18. MEHRA, V., BRENNAN, P. J., RADA, E., CoNvrr, J.and BinoNt, B. R. Lymphocyte suppression inleprosy induced by unique M. leprae glycolipid.Nature 308 (1984) 194-196.
19. MITSUYAMA, N4., loARAsHt, K., KAWAMURA, I.,OrimoRt, T. and Nomo-ro, K. Difference in the in-duction of macrophage interleukin-1 productionbetween viable and killed cells of Listeria mono-cytogenes. Am. Soc. Microbiol. 58 (1990)1254-1260.
20. N4rrstrsANIA, M., WATANABE, M., SANO, K.,AMAKO, K. and NomoTo, K. Generation of Liste- 36.ria monocytogenes-speci tic T cells mediating de-layed foot pad reaction and protection in neona-tally thymectomized mice hut not in nude mice.Med. Microbiol. Immunol. 177 (1988) 207-217.

65, 1^ Yogi, et al.: Cytokine Gene Expression^ 89
tivated macrophages. Proc. Natl. Acad. Sci.U.S.A. 83 (1986) 5233-5237.
45. VA01111,A, M., HoLzER, T. J. and ANDERSEN, B. R.Suppression of monocyte oxidative response byphenolic glycolipid 1 of Mycobacterium /eprae. J.Immunol. 142 (1989) 1696-1701.
46. WONG G. 11. W. and GoiaMEL, D. V. Tumournecrosis factor a and /3 inhibit virus replicationand cynergize with interferons. Nature 323 (1986)819-822.
47. YAMAMURA, M., UYEMURA, K., DEANS, R. J.,WEINBERG, K., RI:A, T. H., BLOOM, B. R. andMODLIN, R. L. Defining protective responses topathogens: cytokine profiles in leprosy lesions.Science 254 (1991) 277-279.
48. YAMAMURA, M., WANG, X.-H., OIIMEN, J. D.,UYEMURA, K., REA, T. H., BLOOM, B. R. and MOD-
LIN, R. L. Cytokine patterns of immunologically
mediated tissue damage. J. Immunol. 149 (1992)1470-1475.
49. Yoca, Y., NAKAMURA, K., Nom:, T., KAWATSU, K.,KASHIWABARA, Y., SAKAMOTO, Y., IZUNII, S.,
SAITO, M., HtoKt, K. and NONURA, T. Susceptibil-ity of severe combined immunodeficient (SCID)mice to Mycobacterium Ieprae: multiplication ofthe bacillus and dissemination of the infection atearly stage. Jpn. J. Lepr. 60 (1991) 139-145.
50. Yoct, Y., NAKAMURA, K. and SUZUKI, A. The ex-perimental inoculation with Mycobacterium lep-rae in autoimmune mice: results of MRL//pr miceinoculated into the right hind foot. Jpn. J. Lepr. 58(1989) 235-240.
51. Yom, Y., NOMAGUCIII, H., MATHUOKA, NI., ET At..
LA study of cytokine mRNAs expression in inocu-lated mice with M. lepraed Proc. Jpn. Soc. 1111-munol. 24 (1994) 554.


















