
COMPARATIVE STUDY OF PREDICTED miRNA BETWEEN INDONESIA AND ...
48
i HALAMAN SAMPUL LAPORAN AKHIR PENELITIAN ILMU KEDOKTERAN COMPARATIVE STUDY OF PREDICTED miRNA BETWEEN INDONESIA AND CHINA (WUHAN) SARS-CoV-2: A BIOINFORMATICS ANALYSIS Tim Pengusul Penelitian Ketua Peneliti : dr Agus Rahmadi, M.Biomed, M.A, NIDN 0426087604 Anggota Peneliti : dr. Ismaily Fasha, M.Ked (ORL-HNS), Sp.THT Nomor Surat Kontrak Penelitian : 318/F.03.07/2020 Nilai Kontrak : Rp. 17.000.000) FAKULTAS KEDOKTERAN PROGRAM PENDIDIKAN DOKTER UNIVERSITAS MUHAMMADIYAH PROF DR HAMKA 2020
Transcript of COMPARATIVE STUDY OF PREDICTED miRNA BETWEEN INDONESIA AND ...

i
HALAMAN SAMPUL
LAPORAN AKHIR
PENELITIAN ILMU KEDOKTERAN
COMPARATIVE STUDY OF PREDICTED miRNA
BETWEEN INDONESIA AND CHINA (WUHAN) SARS-CoV-2:
A BIOINFORMATICS ANALYSIS
Tim Pengusul Penelitian
Ketua Peneliti : dr Agus Rahmadi, M.Biomed, M.A, NIDN 0426087604
Anggota Peneliti : dr. Ismaily Fasha, M.Ked (ORL-HNS), Sp.THT
Nomor Surat Kontrak Penelitian : 318/F.03.07/2020
Nilai Kontrak : Rp. 17.000.000)
FAKULTAS KEDOKTERAN
PROGRAM PENDIDIKAN DOKTER
UNIVERSITAS MUHAMMADIYAH PROF DR HAMKA
2020

ii
HALAMAN PENGESAHAN
PENELITIAN ILMU KEDOKTERAN
Judul Penelitian
STUDI PERBANDINGAN PREDIKSI MIRNA ANTARA SARS-COV-2 INDONESIA
DAN CHINA (WUHAN): TINJAUAN ANALISIS BIOINFORMATIK
Jenis Penelitian : Penelitian Ilmu Kedokteran
Ketua Peneliti : dr Agus Rahmadi, M.Biomed, M.A
Link Profil simakiphttp://simakip.uhamka.ac.id/pengguna/show/1257
Fakultas : Fakultas Kedokteran
Anggota Peneliti : dr. Ismaily Fasha, M.Ked (ORL-HNS), Sp.THT
Link Profil simakip :Click or tap here to enter text.
Contoh link: http://simakip.uhamka.ac.id/pengguna/show/978
Waktu Penelitian : 6 Bulan
Luaran Penelitian
Luaran Wajib : Journal of Infectious Disease
Status Luaran Wajib : In review
Luaran Tambahan : ICNSSE 2020: International Conference On Natural And
Social Science Education
Status Luaran Tambahan: Accepted
Mengetahui, Jakarta, 10 Desember 2020
Ketua Program Studi Ketua Peneliti
dr. Endin Nokik Stujana, PhD dr. Agus Rahmadi, M.Biomed, M.A
NIDN. 0306078805 NIDN.0426087604
Menyetujui,
Dekan Fakultas kedokteran Ketua Lemlitbang UHAMKA
Dr. dr. Wawang Sukarya, SpOG, MARS, MH.Kes Prof. Dr. Suswandari, M.Pd
NIDN. 0030064701 NIDN. 0020116601

iii
SURAT KONTRAK PENELITIAN

iv

v
ABSTRAK
STUDI PERBANDINGAN PREDIKSI miRNA ANTARA SARS-CoV-2 INDONESIA DAN
CHINA (WUHAN) : TINJAUAN ANALISIS BIOINFORMATIK
Agus Rahmadi 1, a), Ismaily Fasyah 1, b)
1Fakultas Kedokteran Universitas Muhamadiyah Prof Hamka, Jakarta 12130, Indonesia
a) Corresponding author: [email protected] b)[email protected]
Abstrak
Beberapa laporan penemuan mutasi dan variasi SARS-CoV-2 pada kasus COVID-19 di
Indonesia menyebabkan disregulasi genom dengan kasus pandemi pertama di Wuhan, China.
MicroRNA (miRNA) memainkan peran penting dalam regulasi genetik dan berkontribusi pada
peningkatan pengikatan RNA virus melalui mRNA inang. Penelitian ini bertujuan untuk mendeteksi
target miRNA SARS-CoV-2 dan mengkaji perannya dalam kasus di Indonesia terhadap kasus di
Wuhan. Secara total, jumlah sampel yang diambil adalah 76 urutan genom lengkap SARS-CoV-2
dari dua kelompok. Perangkat lunak MiRDB digunakan untuk memprediksi target miRNA dengan
referensi miRBase v22.1. Uji chi-square digunakan untuk mendapatkan miRNA yang signifikan
yang membedakan kedua kelompok. Software DIANA-miRPath v3.0 kemudian digunakan untuk
melihat peran target miRNA dalam proses biologis berdasarkan analisis Gene Ontology (GO). Hasil
statistik mendeteksi lima miRNA yang signifikan. Dua miRNA, hsa-miR-4778-5p dan hsa-miR-4531
secara konsisten ditemukan di sebagian besar sampel di Wuhan, sementara hanya kurang dari
setengah sampel di Indonesia. Tiga miRNA lainnya, hsa-miR-6844, hsa-miR-627-5p, dan hsa-miR-
3674, diprediksi pada sebagian besar sampel pada kedua kelompok tetapi dengan rasio yang berbeda
secara signifikan. Di antara lima target miRNA hanya hsa-miR-6844 yang merupakan satu-satunya
miRNA yang terkait dengan gen ORF1ab dari SARS-CoV-2. Ontologi gen yang terdeteksi dari lima
target miRNA menunjukkan peran penting dalam peradangan dan sistem kekebalan selama infeksi
SARS-CoV-2. Deteksi spesifik inang miRNA pada penelitian ini menunjukkan adanya perbedaan
kasus COVID-19 antara di Indonesia dan di Wuhan.
Kata kunci: miRNA, SARS-CoV-2, Indonesia, Wuhan, Bioinformatika

vi
DAFTAR ISI
HALAMAN SAMPUL ....................................................................................................... i
HALAMAN PENGESAHAN ........................................................................................... ii
SURAT KONTRAK PENELITIAN............................................................................... iii
ABSTRAK ......................................................................................................................... v
DAFTAR ISI......................................................................................................................vi
DAFTAR TABEL ............................................................................................................ vii
DAFTAR GAMBAR ....................................................................................................... viii
DAFTAR LAMPIRAN ..................................................................................................... ix
BAB 1. PENDAHULUAN ................................................................................................ 1
BAB 2. TINJAUAN PUSTAKA ....................................................................................... 4
BAB 3. METODE PENELITIAN .................................................................................... 7
BAB 4. HASIL DAN PEMBAHASAN .......................................................................... 10
BAB 5. KESIMPULAN DAN SARAN .......................................................................... 18
BAB 6 LUARAN YANG DICAPAI .............................................................................. 19
BAB VII RENCANA TINDAK LANJUT DAN PROYEKSI HILIRISASI .............. 20
DAFTAR PUSTAKA ...................................................................................................... 21
LAMPIRAN..................................................................................................................... 30

vii
DAFTAR TABEL
Tabel 1. Alur Penelitian
Tabel 2. Karakteristik Sampel Host
Tabel 3. miRNA yang signifikan.
Tabel 4. Tabel Kontingensi miRNA yang signifikan.
Tabel 5. Jalur Ontologi Gen miRNA SARS CoV-2 Indonesia vs Wuhan

viii
DAFTAR GAMBAR
Gambar 1. Proses penelitian
Gambar 2. Skema Jalur prediksi miRNA (has-miR-4778-5p, has-miR-4531, has-
miR-6844) berdasarkan Gen Ontologi dengan kotak kuning sebagai proses biologis
yang dipengaruhi oleh miRNA

ix
DAFTAR LAMPIRAN
Lembar Letter of Acceptance Oral Presentation pada ICNSSE 2020
Lembar Submisson Confirmation pada Journal of Infectious Disease
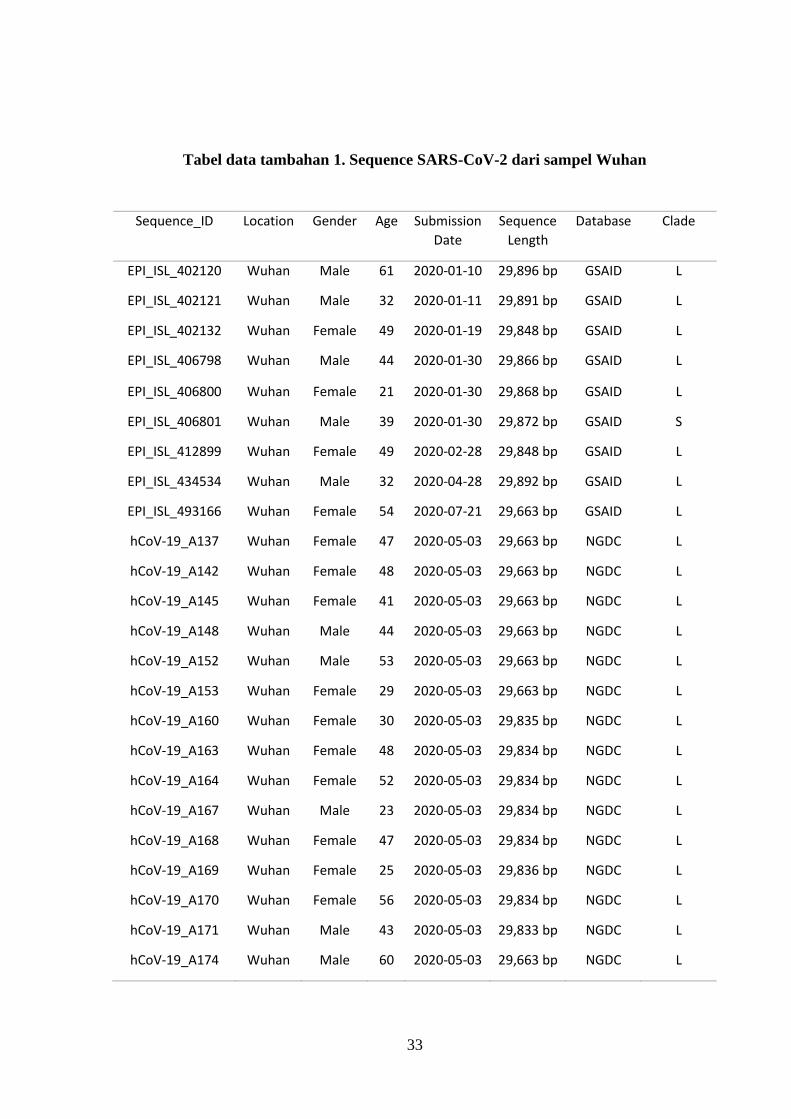
Tabel data tambahan 1. Sequence SARS-CoV-2 dari sampel Wuhan
Tabel data tambahan 2. Sequence SARS-CoV-2 dari sampel Indonesia
Tabel data tambahan 3. Mutasi SARS-CoV-2 dari sampel Wuhan
Tabel data tambahan 4. Mutasi SARS-CoV-2 dari sampel Indonesia

1
BAB 1. PENDAHULUAN
Pandemi virus corona 2019 (COVID-19) yang pertama kali muncul di
Wuhan, Tiongkok pada 12 Desember 2019 disebabkan oleh virus corona baru yang
memiliki kemiripan dengan virus corona kelelawar (kelelawar SL-CoVZC45) pada
tingkat kemiripan nukleotida 89,1% dan berdasarkan pohon filogenetik memiliki
hubungan kuat dengan kelelawar SL-CoVZC45 dan CoVZXC21 dengan nilai
persentase homologi 88% (Wu et al., 2020)(Koyama et al., 2020). Virus yang
dikenal sebagai SARS-CoV-2 (Severe Acute Respiratory Syndrome Coronavirus
2) merupakan keluarga Coronaviridae termasuk virus lain seperti SARS-CoV-1,
dan MERS-CoV dimana sebelumnya menimbulkan KLB pada tahun 2003 dan
2012 (Tu et al., 2020)(de Wit et al., 2016). RNA virus baru ini bermutasi dengan
sangat cepat dan membawa varian baru ke seluruh dunia (Mercatelli & Giorgi,
2020), terdapat 6 varian utama SARS-CoV-2 yang telah diidentifikasi pada 7 Juli
2020. Clade GR telah terdeteksi paling banyak di seluruh dunia (23,4%).
Sedangkan Clade L terdeteksi paling sedikit (6,1%) namun banyak ditemukan pada
kasus pertama COVID-19. Varian yang terdapat di benua Asia sendiri adalah
didominasi oleh Clade O (34,7%) (Hamed S, Elkhatib W, Khairallah A, 2020).
Hingga awal Oktober 2020, Indonesia telah mengkonfirmasi jumlah total
kasus positif COVID-19 adalah 291.182. Persentase kematian di Indonesia adalah
3,72% (10.856 kematian), lebih tinggi dari kasus kematian global 2,98% (WHO,
n.d.). Lonjakan kasus COVID-19 di Indonesia diduga ada kaitannya dengan mutasi
pada gen Spike Glycoprotein (S) yang mengarah pada kemampuan virus untuk
mengikat ACE-2 (Angiotensin-Converting Enzyme 2) reseptor pada manusia
sebagai inang (Hoffmann et al., 2020). Saat ini ada beberapa studi di Indonesia
untuk memahami aspek imunoinformatis (Nidom R V, Indrasari S, Normalina I,
Kusala MKJ, Ansori ANM, 2020)(Ansori et al., 2020), biomolekuler (Arli Aditya
Parikesit & Nurdiansyah, 2020)(Arli Ardi Parikesit & Nurdiansyah, 2020), varian
genetik, dan mutasi (Nidom R V, Indrasari S, Normalina I, Kusala MKJ, Ansori
ANM, 2020)(Muhammad Ansori et al., 2020) genom SARS-CoV-2 dari Indonesia.
Namun penelitian tentang microRNA (miRNA) SARS-CoV-2 masih jarang

2
dilakukan di Indonesia. Oleh karena itu, kami melakukan prediksi target dan
profiling miRNA dari sampel Indonesia dan membandingkannya dengan sampel
dari Wuhan.
MicroRNA adalah RNA non-coding yang memiliki peran penting dalam
regulasi dan ekspresi genetic sebuah organisme. Disregulasi mikroRNA dapat
menyebabkan perubahan pola ekspresi dan mutasi gen yang menyebabkan
keparahan penyakit (Fernandez et al., 2017)(Winter et al., 2009)(Krol et al.,
2010)(Mendell & Olson, 2012)(Finnegan & Pasquinelli, 2013). Selain itu,
perubahan fungsi miRNA dapat dipengaruhi oleh variasi genetik yang dapat
diidentifikasi dengan mendeteksi miRNA target dan urutan miRNA (Fernandez et
al., 2017)(Cammaerts et al., 2015)(Martin-Guerrero et al., 2015). Dalam beberapa
kasus, patogenisitas dan replikasi RNA virus dimediasi oleh pengikatan langsung
miRNA inang menjadi RNA virus yang menginduksi disregulasi genetik pada inang.
RNA virus mempengaruhi jalur regulasi host dengan meniru miRNA seluler yang
secara langsung mempengaruhi transkriptom host (Skalsky & Cullen,
2010)(Trobaugh & Klimstra, 2017). Peran miRNA terhadap penyakit infeksi salah
satunya SARS-CoV-2 merupakan topik yang menarik untuk diteliti sehingga
banyak jurnal yang telah diterbitkan sejak wabah COVID-19 pada awal 2020.
Beberapa penelitian juga mencoba mempelajari sampel dan metode yang
berbeda dalam memprediksi miRNA, salah satunya adalah studi oleh Liu dan rekan
(Liu et al., 2020). Mereka menggunakan perangkat lunak ab initio yang disebut
VMir (Grundhoff, 2011) untuk mengidentifikasi beberapa potensi pra-miRNA dari
genom SARS-CoV-2 dengan nomor aksesi MN908947. MatureBayes (Gkirtzou et
al., 2010) digunakan juga untuk memprediksikan miRNA. Selain itu target miRNA
pada virus dan genom manusia juga diprediksi menggunakan TargetFinder
(miRanda). Lebih lanjut, database Target Scan Human 7.2 digunakan untuk
menemukan gen manusia yang menjadi target yang miRNA. Gen Ontologi (GO)
digunakan untuk menjelaskan secara fungsional gen terkait.
Studi lain juga menggunakan VMIR untuk menentukan kandidat pra-
miRNA dari genom SARS-CoV-2 (Sarma et al., 2020). Kandidat pra-miRNAs dicari

3
kemiripan nukleotida nya dengan miRNA manusia yang tersedia dalam database
miRbase. Alat hybrid RNA paling banyak digunakan untuk hibridisasi antara pra-
mirRNA virus dan miRNA manusia. Identifikasi miRNAs menggunakan
MatureBayes. Untuk selanjutnya Analisis GO dari gen target dilakukan oleh
PANTHER (Protein Analysis Through Evolutionary Relationships). Demirci dan
Adan (Saçar Demirci & Adan, 2020) menggunakan izMiR untuk memprediksi pra-
miRNA dari miRNA jepit rambut, mereka memanfaatkan psRNATarget untuk
mencari miRNA yang ditargetkan.
Selanjutnya, miRBase digunakan untuk mencari target dalam gen SARS-
CoV-2. Namun, penelitian lain langsung mencari miRNA tanpa menentukan pra-
miRNA, yang dapat dilakukan dengan menggunakan perangkat lunak miRDB
berbasis web, terutama miRNA yang memengaruhi regulasi gen secara langsung
(Fulzele et al., 2020)(Haddad & Walid Al-Zyoud, 2020). Fulzele dan rekan (Fulzele
et al., 2020), mengamati peran miRNA dengan gen fungsional manusia melalui
analisis jalur KEGG dan analisis GO pada perangkat lunak DIANA-miRPath v.3.0
berdasarkan skor prediksi target di atas 90 dari miRDB.
Dalam penelitian ini, kami menggunakan sampel SARS-CoV-2 yang
diperoleh dari Indonesia dan membandingkannya dengan sampel Wuhan untuk
memprediksi target miRNA dengan menggunakan analisis bioinformatika.
Penelitian ini bertujuan untuk menemukan miRNA inang target SARS-CoV-2 dan
peran patogenesisnya dalam kasus COVID-19.

4
BAB 2. TINJAUAN PUSTAKA
Kasus COVID-19 terjadi di setidaknya 170 negara dengan tingkat kematian
sekitar 5,4%. Secara umum, sebagian besar kasus berusia 30-79 tahun (86,6%),
dengan kelompok usia ≥80 ditandai dengan tingkat kematian tertinggi di antara
semua kelompok umur (Chen et al., 2020). Di Indonesia sendiri, berdasarkan situs
resmi gugus tugas percepatan penanganan covid 19 pada tanggal 29 April 2020
dilaporkan jumlah kasus positif adalah sebanyak 9771 orang. (Gugus Tugas
Percepatan Penanganan Covid 19, n.d.) Gejala COVID-19 diantaranya demam
(83-98%), batuk (59-82%), sesak napas (19-55%), dan nyeri otot (11-44%), yang
mirip dengan SARS (Severe Acute Respiratory Syndrome) dan MERS (Middle East
Respiratory Syndrome) (Huang et al., 2020). Beberapa pasien mungkin mengalami
sakit tenggorokan, sakit kepala, dan sebagian kecil pasien mengalami hemoptisis
(Huang et al., 2020)(Guan et al., 2020) dan sejumlah kasus ditemukan relatif tanpa
gejala (Chan et al., 2020). Masa inkubasi virus rata-rata 5-6 hari dengan masa
inkubasi terpanjang 14 hari. Pada kasus COVID-19 yang berat dapat menyebabkan
pneumonia, sindrom pernapasan akut, gagal ginjal, dan bahkan kematian.
Berdasarkan bukti ilmiah, COVID-19 dapat menular dari manusia ke manusia
melalui percikan batuk/bersin (droplet), tidak melalui udara (Pedoman Pencegahan
Dan Pengendalian Corona Virus Disease (COVID - 19), 2020).
SARS-CoV, MERS-CoV, dan SARS-CoV-2 merupakan keluarga
Coronaviridae. Kelompok virus ini mengandung genom RNA berantai positif,
dengan untai tunggal yang relatif besar, sekitar 27-32 kb (Pyrc et al., 2006). Dengan
Genom yang terdiri dari cap 50-metilguanosin di awal, tail 30-poli-A di akhir,
dengan total 6-10 gen (de Wit et al., 2016). Di sisi lain, protein struktural, termasuk
Spike (S), Envelope (E), dan Membrane (M) merupakan mantel virus, dan protein
Nukleokapsid (N) yang mengemas genom virus (RNA). Beberapa protein ini
mengalami glikosilasi dalam Aparatus Golgi untuk membentuk glikoprotein. Di
antara semua protein struktural, target terapi potensial yang paling penting adalah
spike (S) glikoprotein, yang bertanggung jawab untuk pengikatan virus ke sel inang.
Protein S dipersiapkan oleh protease sel inang dan dikenali oleh reseptor seluler.

5
Protein serine manusia yaitu Transmembrane Serine Protease 2 (TMPRSS2)
bertanggung jawab untuk menentukan protein S dari SARS-CoV dan SARS-CoV-2,
dan Angiotensin Converting Enzym 2 (ACE2) yang terlibat sebagai reseptor untuk
masuknya kedua virus ini. Sedangkan untuk MERS-CoV, ia berikatan dengan
reseptor lain, Di Peptidyl Peptidase 4 (DPP4) (Hoffmann et al., 2020). Perlu dicatat
bahwa tingkat ekspresi ACE2, dan juga frekuensi alel bervariasi di antara populasi
(Cao et al., 2020). Karakteristik reseptor seluler ACE2 juga dapat menjelaskan
patogenesis dari SARS-CoV dan SARS-CoV-2. Seperti diketahui bahwa pengikatan
protein S virus ke ACE2 menginduksi umpan balik negatif yang akhirnya
menghasilkan downregulasi ACE2. Penurunan ACE2 selanjutnya menyebabkan
permeabilitas pembuluh darah paru meningkat (Kuba et al., 2005).
MicroRNA (miRNA) merupakan keluarga RNA yang tidak menyandi (non–
coding RNA) yang tersusun atas 19–24 nukleotida yang menghambat translasi
mRNA dan meningkatkan degradasi mRNA, hasilnya akan menurunkan tingkat
ekspresi protein. miRNA secara khusus mengikatkan diri pada regio 3’ dari mRNA
yang tidak tertranslasi (untranslated region) dan menyebabkan represi dan atau
degradasi paska transkripsi mRNA bersangkutan dalam sel yang berproliferasi.
MiRNA terletak dalam intron maupun ekson dari gen penyandi protein (70%) atau
di daerah intergenik (30%). MiRNA memiliki peran biologis untuk mengatur
berbagai proses penting dalam pertumbuhan, diferensiasi, apoptosis, proliferasi,
penuaan dan proses seluler lain. Penelitian bioinformatika dan microarray
mengungkapkan bahwa satu miRNA tunggal dapat berikatan dengan 200 gen
sasaran, dan bahwa gen sasaran ini dapat berupa faktor transkripsi, reseptor, faktor
yang disekresikan maupun transporter (Xia HP., 2008).
MiRNA yang disandikan pada virus pertama kali ditemukan pada Epstein-
Barr Virus (EBV) manusia (Pfeffer, 2004) dan lebih dari 320 prekursor miRNA virus
dilaporkan sejauh ini. Seperti diketahui bahwa berbagai virus DNA dapat
mengekspresikan miRNA, namun bagaimana dengan virus RNA apakah bisa
mengekspresikan miRNA, hal ini masih dalam perdebatan, karena virus RNA

6
bereplikasi dalam sitoplasma dan tidak memiliki akses ke mesin miRNA di inti sel
(Demirci et al., 2016).
ROADMAP PENELITIAN
BATCH I (2020)
• Identifikasi micro RNA SARS-COV-2
• (Penelitian yang dilakukan)
BATCH II (2021)
• Pemeriksaan micro RNA yang berperan dalam SARS-COV-2 berdasarkan identifikasi yang di dapat pada BATCH I
BATCH III (2022)
• Target pengobatan dengan micro RNA pada uji pra klinik

7
BAB 3. METODE PENELITIAN
DATA GENOM SEQUENCE SARS COV-2
Data genom sequence SARS CoV-2 diperoleh dari 37 genom lengkap dari
GISAID (https://www.gisaid.org/), NCBI (https://ncbi.nlm.nih.gov), dan National
Genomics Data Center (NGDC) yang merupakan bagian dari China National
Center for Bioinformation (https://bigd.big.ac.cn/gwh/) sebagai referensi genom
(tabel data tambahan 1). Selain itu, kami memperoleh 38 Data genom sequence
SARS CoV-2 di Indonesia dari GISAID yang diakses pada 13 September 2020
(tabel data tambahan 2).
PREDIKSI MIRNA
Perangkat lunak MiRDB digunakan untuk memprediksi miRNA terhadap
urutan SARS CoV-2. Perangkat lunak miRDB menggunakan kode yang dijalankan
dari gbnegrini/mirdb-custom-target-search (https://github.com/gbnegrini/mirdb-
custom-target-search) berdasarkan versi perangkat lunak berbasis web (http://
mirdb.org/custom.html). Dalam pekerjaan ini, kami menggunakan algoritma
mirTarget berdasarkan Support Vector Machine untuk menghitung skor prediksi
target miRNA (X. Wang, 2008). Ambang batas skor prediksi target miRNA yang
digunakan dalam pekerjaan kami adalah 80. Target miRNA yang terdeteksi oleh
miRDB telah cocok berdasarkan jumlah miRNA dari database miRBase versi 22.1
(http://www.mirbase.org/). Selain itu, kami juga menggunakan perangkat lunak
ChromosPros Versi 2.1.9 untuk quality control sequence SARS CoV-2.
ANALISIS STATISTIK
Untuk mendapatkan miRNA yang signifikan dari kedua kelompok, maka
dilakukan serangkaian uji Chi-Square untuk melihat perbedaan dengan melihat
proporsi tingkat kemunculan miRNA pada kedua kelompok. Hanya miRNA dengan
nilai p <0,05 dianggap signifikan.

8
ANALISIS JALUR MIRNA
miRNA yang signifikan dianalisis jalur mereka dengan menggunakan perangkat
lunak berbasis web DIANA-miRPath v 3.0 (http://www.microrna.gr/miRPathv3)
yang berbasis Gene Ontology (GO) dengan subkategori proses biologis (Vlachos et
al., 2015). DIANA-miRPath v3.0 menerapkan algoritma DIANA-microT-CDS
yang memprediksi CDS (Coding Sequence) mRNA dan 3 'UTR region dengan
akurasi tinggi (Paraskevopoulou et al., 2013). Keseluruhan proses penelitian dapat
dilihat pada Gambar 1.
Gambar 1. proses penelitian
9
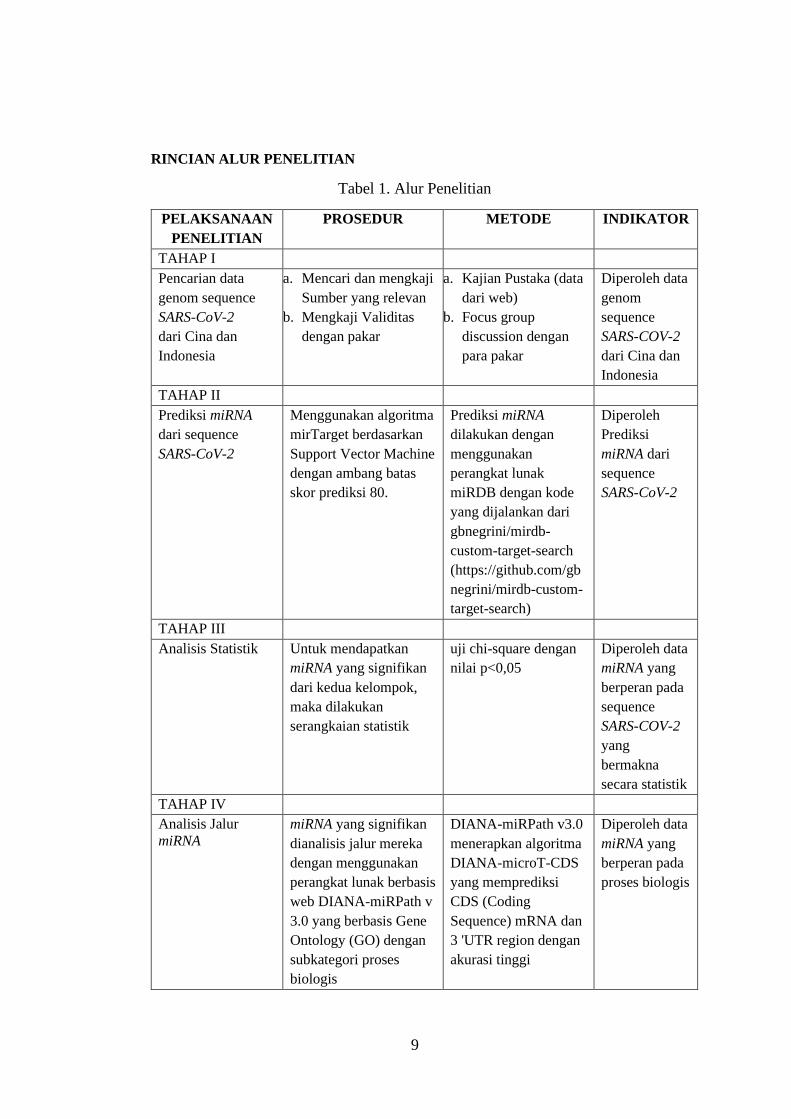
RINCIAN ALUR PENELITIAN
Tabel 1. Alur Penelitian
PELAKSANAAN
PENELITIAN
PROSEDUR METODE INDIKATOR
TAHAP I
Pencarian data
genom sequence
SARS-CoV-2
dari Cina dan
Indonesia
a. Mencari dan mengkaji
Sumber yang relevan
b. Mengkaji Validitas
dengan pakar
a. Kajian Pustaka (data
dari web)
b. Focus group
discussion dengan
para pakar
Diperoleh data
genom
sequence
SARS-COV-2
dari Cina dan
Indonesia
TAHAP II
Prediksi miRNA
dari sequence
SARS-CoV-2
Menggunakan algoritma
mirTarget berdasarkan
Support Vector Machine
dengan ambang batas
skor prediksi 80.
Prediksi miRNA
dilakukan dengan
menggunakan
perangkat lunak
miRDB dengan kode
yang dijalankan dari
gbnegrini/mirdb-
custom-target-search
(https://github.com/gb
negrini/mirdb-custom-
target-search)
Diperoleh
Prediksi
miRNA dari
sequence
SARS-CoV-2
TAHAP III
Analisis Statistik Untuk mendapatkan
miRNA yang signifikan
dari kedua kelompok,
maka dilakukan
serangkaian statistik
uji chi-square dengan
nilai p<0,05
Diperoleh data
miRNA yang
berperan pada
sequence
SARS-COV-2
yang
bermakna
secara statistik
TAHAP IV
Analisis Jalur
miRNA
miRNA yang signifikan
dianalisis jalur mereka
dengan menggunakan
perangkat lunak berbasis
web DIANA-miRPath v
3.0 yang berbasis Gene
Ontology (GO) dengan
subkategori proses
biologis
DIANA-miRPath v3.0
menerapkan algoritma
DIANA-microT-CDS
yang memprediksi
CDS (Coding
Sequence) mRNA dan
3 'UTR region dengan
akurasi tinggi
Diperoleh data
miRNA yang
berperan pada
proses biologis

10
BAB 4. HASIL DAN PEMBAHASAN
HASIL
Secara total, ada 76 sampel genom SARS-COV-2 yang dimasukkan dalam
penelitian ini. Rangkuman karakteristik sampel host tersebut berdasarkan jenis
kelamin dan umur dapat dilihat pada Tabel 2. Sampel host Indonesia sebagian besar
berjenis kelamin laki-laki. Sebaliknya, sampel Wuhan mayoritas berjenis kelamin
perempuan. Tidak ada perbedaan usia yang signifikan dari kedua sampel tersebut.
Tabel 2. Karakteristik Sampel Host
Indonesia Wuhan P-value
Gender 0.011
Male 29 (74.36%) 17 (45.95%)
Female 10 (25.64%) 20 (54.05%)
Age 0.061
range 17-83 21-65
mean 50.79 44.19
stdev 17.89 11.56
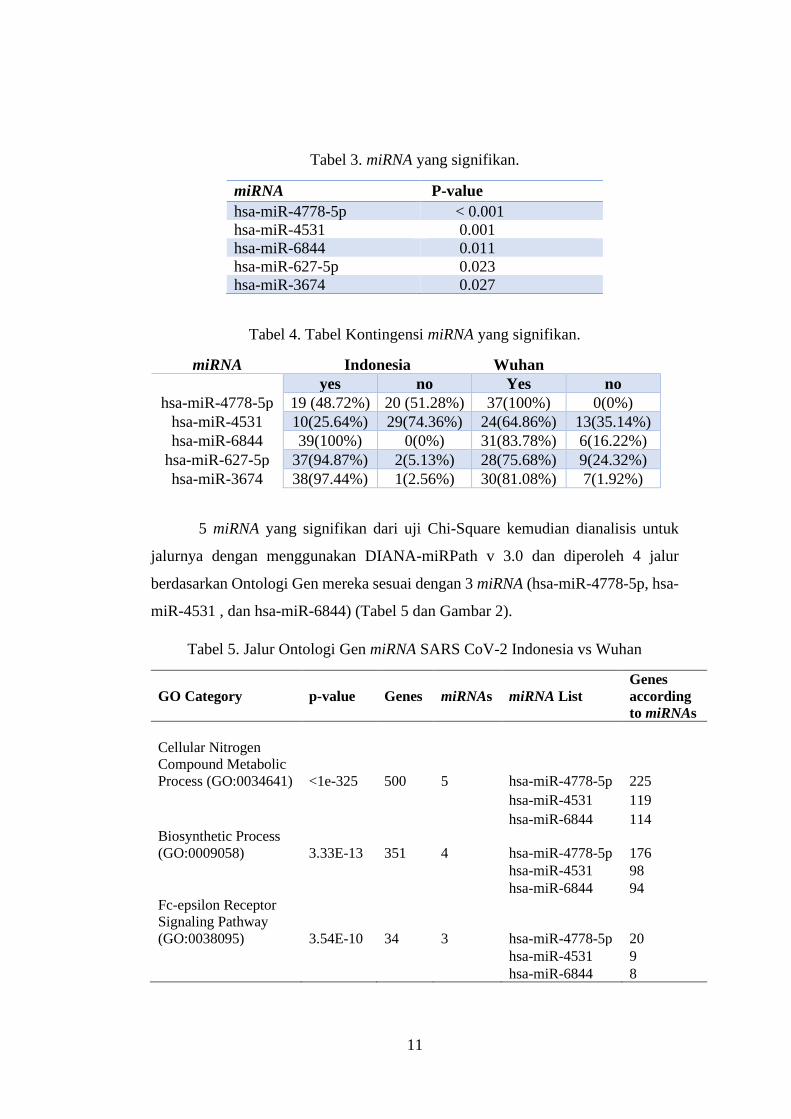
Hasil Uji Chi-Square menunjukkan lima miRNA yang signifikan pada kedua
kelompok ini. Tabel 3 menunjukkan daftar miRNA yang signifikan dengan nilai p
< 0,05. Tabel kontingensi lengkap untuk miRNA ini dapat dilihat pada Tabel 4. hsa-
miR-4778-5p dan hsa-miR-4531 adalah dua miRNA yang secara jelas terlihat
kontras antara kedua kelompok ini. Kedua miRNA ini tidak diprediksi pada lebih
dari 50% sampel genom dari Indonesia tetapi diprediksi sebagian besar sampel
Wuhan. Selain itu, has-miR-4788-5p secara konsisten diprediksi di semua sampel
Wuhan. Tiga miRNA lainnya diprediksi pada kedua kelompok sampel meskipun
dengan persentase yang berbeda. Di antara kelima miRNA tersebut, hanya
memiliki-miR-6844 yang berasosiasi dengan gen ORF1AB.
11
Tabel 3. miRNA yang signifikan.
miRNA P-value
hsa-miR-4778-5p < 0.001
hsa-miR-4531 0.001
hsa-miR-6844 0.011
hsa-miR-627-5p 0.023
hsa-miR-3674 0.027
Tabel 4. Tabel Kontingensi miRNA yang signifikan.
miRNA Indonesia Wuhan
yes no Yes no
hsa-miR-4778-5p 19 (48.72%) 20 (51.28%) 37(100%) 0(0%)
hsa-miR-4531 10(25.64%) 29(74.36%) 24(64.86%) 13(35.14%)
hsa-miR-6844 39(100%) 0(0%) 31(83.78%) 6(16.22%)
hsa-miR-627-5p 37(94.87%) 2(5.13%) 28(75.68%) 9(24.32%)
hsa-miR-3674 38(97.44%) 1(2.56%) 30(81.08%) 7(1.92%)
5 miRNA yang signifikan dari uji Chi-Square kemudian dianalisis untuk
jalurnya dengan menggunakan DIANA-miRPath v 3.0 dan diperoleh 4 jalur
berdasarkan Ontologi Gen mereka sesuai dengan 3 miRNA (hsa-miR-4778-5p, hsa-
miR-4531 , dan hsa-miR-6844) (Tabel 5 dan Gambar 2).
Tabel 5. Jalur Ontologi Gen miRNA SARS CoV-2 Indonesia vs Wuhan
GO Category p-value Genes miRNAs miRNA List
Genes
according
to miRNAs
Cellular Nitrogen
Compound Metabolic
Process (GO:0034641) <1e-325 500 5 hsa-miR-4778-5p 225
hsa-miR-4531 119
hsa-miR-6844 114
Biosynthetic Process
(GO:0009058) 3.33E-13 351 4 hsa-miR-4778-5p 176
hsa-miR-4531 98
hsa-miR-6844 94
Fc-epsilon Receptor
Signaling Pathway
(GO:0038095) 3.54E-10 34 3 hsa-miR-4778-5p 20
hsa-miR-4531 9
hsa-miR-6844 8

12
Cellular Protein
Modification Process
(GO:0006464) 6.49E-10 220 3 hsa-miR-4778-5p 106
hsa-miR-4531 61
hsa-miR-6844 68
Gambar 2. Skema Jalur prediksi miRNA (hsa-miR-4778-5p, hsa-miR-4531, hsa-
miR-6844) berdasarkan Gen Ontologi dengan kotak kuning sebagai proses
biologis yang dipengaruhi oleh miRNA
PEMBAHASAN
Lima miRNA yang tercantum dalam Tabel 1 merupakan miRNA yang
termasuk dalam daftar 873 miRNA umum yang menargetkan genom COVID-19
yang ditemukan secara analisis in silico oleh Fulzele dan rekan (Fulzele et al.,
2020). Mereka menggunakan 29 isolat COVID-19 dari berbagai negara, termasuk
dari Wuhan dan negara Asia lainnya. Selain itu, temuan mereka menunjukkan
bahwa lima miRNA kami juga termasuk dalam 558 isolat COVID-19 yang
umumnya ada. Namun, kami tidak menemukan satupun dari lima miRNA dengan
skor target tertinggi (di atas 94) dari perhitungan kami cocok dengan hasil mereka

13
(7 miRNA). Selain itu, Khan dan rekan (Khan et al., 2020) mencoba mempelajari
perbedaan pola pengikatan miRNA inang dalam berbagai genom COVID-19 (67
isolat) dari wilayah yang berbeda (24 negara). Mereka menemukan 24 miRNA
inang dan melakukan analisis clustering untuk menemukan dua cluster miRNA
utama yang berasal dari isolat Asia (Isolat Korea Selatan dan Singapura). Mirip
dengan hasil Fulzele dan rekan, tetapi penelitian kami tidak menemukan satupun
prediksi miRNA dengan miRNA pada penelitian Khan dan rekan, sehingga hasil
kami yang unik dan berbeda memerlukan penyelidikan lebih lanjut. Selain itu, studi
terbaru oleh Li dan rekan mengamati perbedaan ekspresi miRNA dalam darah tepi
antara pasien COVID-19 dan donor sehat, mereka menemukan miR ‐16-2‐3p, miR‐
6501‐5p, dan miR‐618 secara signifikan diekspresikan pada pasien COVID-19
dibandingkan dengan donor yang sehat (Li et al., 2020).
Selain COVID-19, miRNA yang kami temukan ternyata memiliki kaitan
dengan tumor berdasarkan penelitian sebelumnya. Hal ini dapat menjadi
pertimbangan untuk mengembangkan strategi dalam menangani penyebaran
SARS-CoV-2. Misalnya, hsa-miR4778, dilaporkan memiliki hubungan dengan
tumor di jaringan payudara (Persson et al., 2011). Pasien yang terinfeksi SARS-
CoV-2 di Wuhan memiliki miRNA ini lebih banyak daripada di Indonesia, hal ini
mungkin disebabkan SARS-CoV-2 di Wuhan menginfeksi orang yang sudah
memiliki tumor di jaringan payudara atau mungkin menyebabkan tumor di jaringan
payudara. Di sisi lain, hsa-miR-627 yang terdeteksi pada pasien yang terinfeksi
SARS-CoV-2 di Indonesia lebih banyak dibandingkan pasien di Wuhan yang
memiliki keterkaitan dengan kanker kolorektal (Cummins et al., 2006). Selain itu
hsa-miR-3674 juga sering ditemukan di pasien Indonesia, dan berkaitan dengan
tumor (Vaz et al., 2010). Selain tumor, hsa-miR-4531 memiliki hubungan dengan
keganasan sel B pada manusia (Jima et al., 2010). Terakhir, hsa-miR-6844 juga
ditemukan pada tikus (Bickels et al., 2001). Hal ini mungkin menunjukkan bahwa
SARSCoV-2 dapat menyebar ke tikus dan menjadi pembawa. Aspek ini harus
diteliti lebih lanjut.

14
Hasil analisis prediksi target miRNA kami menunjukkan sedikit perbedaan
antara SARSCoV-2 yang ditemukan di Indonesia dan Wuhan. Perbedaan ini
mungkin disebabkan oleh mutasi (perubahan genetika minor) atau rekombinasi
(perubahan genetika mayor) (Wimmer & Goldbach, 1992). Namun dalam studi
pendahuluan ini, kami hanya memfokuskan pembahasan kami pada mutasi-mutasi
yang menyebabkan perbedaan SARS-CoV-2 dari kedua sumber ini. Tabel data
tambahan 3 dan 4 menunjukkan mutasi yang ditemukan di setiap sampel dari kedua
sumber. Dalam sampel Indonesia, tiga besar mutasi yang paling umum adalah Spike
D614G, NSP12 P323L, dan N3 Q57H. Ketiga Mutasi tersebut tidak identik dengan
tiga mutasi paling umum pada sampel Wuhan, yaitu NSP2 N92H, Spike N856K,
dan NS8 L84S. Pengaruh mutasi Spike D614G berhubungan dengan infeksiusitas
SARS-CoV-2 dan berhungan dengan pandemic global (Mercatelli & Giorgi,
2020)(Korber et al., 2020)(Plante et al., 2020)(Mohammad et al., 2020)(Jackson et
al., 2020)(Yurkovetskiy et al., 2020)(Franco-Muñoz et al., 2020). Mutasi ini sangat
terkait dengan tingkat infeksi yang lebih tinggi pada sel paru-paru manusia dan sel
usus besar (Yurkovetskiy et al., 2020). Sebuah studi dari Amerika Selatan
menemukan bahwa mutasi khusus ini terjadi pada 83% sampel mereka dan
menyarankan investigasi lebih lanjut terkait dengan perkembangan vaksin dan
metode pengobatan lainnya.
Selain itu, kombinasi Spike D614G dan NSP12 P323L (mutasi umum
pertama dan kedua pada sampel Indonesia) pada semua varian SARS-CoV-2 yang
terdeteksi menunjukkan pentingnya dua mutasi ini dalam hal penularan dan tingkat
patogenisitas (Mutlu et al., 2020)(Hartley et al., 2020)(Kannan et al., 2020). Sebuah
studi dari kasus Vietnam, juga menemukan D614G dan P323L merupakan mutasi
yang paling umum, begitu juga di eropa di dapatkan hal yang sama (Nguyen et al.,
2020). Berbeda dengan kedua mutasa sebelumnya, terdapat juga sedikit penelitian
yang membahas mutasi N3 Q57H (R. Wang et al., 2020)(Nguyen et al.,
2020)(Soratto et al., 2020)(Soratto et al., 2020)(Hassan et al., 2020). Namun mutasi
ini ditemukan di 70% sampel Amerika Serikat bersama dengan mutasi lain yaitu
T85I (R. Wang et al., 2020). Studi tersebut juga menunjukkan bahwa mutasi ini

15
pertama kali ditemukan di Singapura pada pertengahan Februari 2020 (R. Wang et
al., 2020).
Selain faktor mutasi, kami juga menyertakan analisis clade untuk
membandingkan karakteristik SARS-CoV-2 dari Indonesia dan Wuhan. Clade
dapat dikaitkan dengan analisis filogenetik yang mengelompokkan varian virus
tertentu berdasarkan garis keturunan. Antara Sampel SARS-CoV-2 Indonesia
sebagian besar dikelompokkan ke dalam clade GH (50%) dan L (40%), dengan
sisanya, dikategorikan ke dalam klade G, O, dan GR. Di sisi lain, semua sampel
dari Wuhan dikelompokkan dalam clade L kecuali satu sampel yang
dikelompokkan dalam clade S. Clade L secara dominan diwakili dalam sampel Asia
(Mercatelli & Giorgi, 2020). Hal ini menyebabkan mayoritas sampel termasuk
sampel Wuhan dan hampir separuh Indonesia juga dikelompokkan dalam clade L.
Menariknya, clade GH sebagai kelompok sampel yang paling umum di Indonesia,
sebagian besar telah terdeteksi di Benua Amerika (Mercatelli & Giorgi, 2020)(B.
Hu et al., 2020). Clade ini berasal dari mutasi D614G dan Q57H yang biasa
ditemukan dalam sampel Indonesia. Saat ini, clade GH bersama GR adalah clade
yang paling umum diamati di semua sampel sekuensing SARS-CoV-2 secara global
(Mercatelli & Giorgi, 2020). Dari analisis, jumlah clade GH sangat banyak
jumlahnya pada Mei 2020 (Alm et al., 2020).
Dari sisi analisis ontologi gen, terdapat 4 jalur yang menunjukkan peran hsa-
miR-4778-5p, hsa-miR-4531, dan hsa-miR-6844 dalam proses biologis (Tabel 5).
Cellular Protein Modification Process (GO: 0006464) mendeteksi target miRNA
yang berperan dalam Post Translational Modification (PTM). Proses glikosilasi
merupakan salah satu PTM dalam SARSCoV-2 dan ditemukan pada sebagian besar
kompleks N-glikan dalam protein S dan ACE2 (Angiotensin Converting Enzyme
2) pada manusia menurut sebuah studi Sun dan rekan (Sun et al., 2020). Sementara
itu, Bouhaddo dan rekan (Bouhaddou et al., 2020) meneliti fosfoproteomik pada
studi "in vivo" dan "in silico" yang menunjukkan penurunan proses fosforilasi pada
NSP12 (Non Structural Protein 12) selama infeksi SARS-CoV-2. Penurunan
fosforilasi ini dapat dikaitkan dengan peningkatan stres oksidatif yang

16
mengoksidasi protein untuk menginduksi mutasi (Valko et al., 2007). Pada sampel
Indonesia telah terjadi mutasi NSP12 sebanyak 43 kali, hal ini memungkinkan
peran miRNA terhadap PTM pada regulasi protein fungsional selama infeksi
SARS-CoV-2. Protein non-struktural ini menunjukkan korelasi yang kuat pada
target miRNA yang terkait dengan gen ORF1ab.
Cellular Nitrogen Compound Metabolic Process (GO: 0034641) dan
Biosynthetic Process (GO: 0009058) memiliki korelasi yang kuat (lihat gambar 2)
berdasarkan 3 target miRNA untuk SARSCoV-2 yang terdeteksi. Kedua proses
biologis tersebut menunjukkan peran pada proses inflamasi selama infeksi SARS-
CoV-2. Studi molekuler SARS-CoV-2 yang dilakukan oleh Hu dan rekan (X. Hu
et al., 2020) menunjukkan peran yang sama dalam proses inflamasi pada paru-paru.
Penelitian Arisan dan rekan (Arisan et al., 2020) juga mendeteksi Cellular Nitrogen
Compound Metabolic Process berdasarkan analisis Gen Ontologi pada sekuens
genom dari Cina, Turki, Italia, dan Spanyol. Proses biosintetik yang terkait dengan
SARS-CoV-2 dalam proses inflamasi sebagian besar terkait dengan biosintesis
asam lemak, terutama asam lemak arakidonat dan asam lemak tak jenuh (Abu-
Farha et al., 2020)(Geurdes & Koutsaroff, 2020)(Kothapalli et al., 2020)(Shawky
et al., 2020). Dalam jalur biosintesis arakidonat, komponen pro-inflamasi dan
produk senyawa endogen diproduksi dan menyebabkan peradangan. Probabilitas
infeksi SARS-CoV-2 bisa meningkat dengan defisiensi regulasi jalur asam
arakidonat (Das, 2020)(Hoxha, 2020). Selain biosintesis asam lemak, Cellular
Protein Modification Process, Cellular Nitrogen Compound Metabolic Process, dan
Biosynthetic Process berperan juga dalam disregulasi metabolisme nitrogen melalui
perubahan komponen dan biosintesis asam amino yang menunjukkan peningkatan
biomarker stres oksidatif selama infeksi SARS-CoV-2 (Thomas et al., 2020).
Analisis Gen Ontologi lainnya adalah Fc-epsilon Receptor Signaling Pathway (GO:
0038095), yang memiliki peran kunci dalam reaksi imunologi yang terkait dengan
SARSCoV-2. Menurut sebuah studi oleh Fulzele dan rekan (Fulzele et al., 2020),
terdapat 24 target miRNA host pada infeksi SARS-CoV2 yang berperan dalam Fc-
epsilon Receptor Signaling Pathway. Sebuah studi pendahuluan proteomik oleh
Leng dan rekan (Leng et al., 2020) dengan sampel klinis terkait cedera paru-paru

17
yang disebabkan oleh COVID-19, menunjukkan penurunan regulasi beberapa
protein yang terkandung dalam Fc-epsilon Receptor Signaling Pathway. Perubahan
regulasi pada cedera paru terkait COVID-19 juga diikuti oleh beberapa sinyal
terkait imun lainnya seperti reseptor lektin tipe-C dan NF-kb. Efek dari Fc-epsilon
Receptor Signaling Pathway juga ditemukan dalam studi Ouyang dan rekan
(Ouyang et al., 2020), dimana study tersebut mendeteksi ekspresi berlebih dari
MAP2K7 dan SOS1 yang mempengaruhi jalur pensinyalan reseptor sel-T yang
diperkirakan berperan dalam perkembangan COVID-19. Fc-epsilon Receptor
Signaling Pathway terkait dengan Cellular Protein Modification Process yang
berhubungan dengan mekanisme fosforilasi / defosforilasi dalam reseptor IgE
dengan afinitas tinggi yang disebut Fc epsilon RI (FcεRI). Selain itu, reaksi Fc
epsilon RI terhadap antigen IgE juga berperan dalam proses inflamasi dengan
melepaskan mediator inflamasi (Paolini et al., 1992)(Siraganian, 2003)

18
BAB 5. KESIMPULAN DAN SARAN
Deteksi spesifik miRNA pada penelitian ini menunjukkan adanya
perbedaan kasus COVID-19 di Indonesia dan China (Wuhan), khususnya dalam
studi biologi molekuler. Lima miRNA inang target yang signifikan menunjukkan
peran penting dalam peradangan dan sistem kekebalan selama infeksi SARS-CoV-
2. Perlu dilakukan penelitian lebih lanjut terhadap lima miRNA dengan targetnya
yang dapat dimanfaatkan untuk biomarker potensial atau untuk tujuan pengobatan
pada kasus COVID-19 di Indonesia.

19
BAB 6 LUARAN YANG DICAPAI
Luaran yang dicapai berisi Identitas luaran penelitian yang dicapai oleh
peneliti sesuai dengan skema penelitian yang dipilih.
Jurnal
IDENTITAS JURNAL
1 Nama Jurnal Journal of Infectious Diseases
2 Website Jurnal https://www.tandfonline.com/toc/infd20/current
3 Status Makalah Review
4 Jenis Jurnal Jurnal International
4 Tanggal Submit 30-Nov-2020
5 Bukti Screenshot submit
Pemakalah di seminar
IDENTITAS SEMINAR
1 Nama Jurnal ICNSSE 2020: International Conference On Natural
And Social Science Education
2 Website Jurnal https://conference.uhamka.ac.id/lic
3 Status Makalah Accepted
4 Jenis Prosiding Prosiding International
4 Tanggal Submit 09 November 2020
5 Bukti Screenshot submit
Pemakalah di seminar

20
BAB VII RENCANA TINDAK LANJUT DAN PROYEKSI HILIRISASI
Minimal mencakup 2 hal ini.
Hasil Penelitian Seperti kita ketahui bahwa penyakit yang disebabkan oleh
virus SARS-CoV-2 masih terus meningkat dari hari ke hari,
sedangkan pengobatan masih terus dikembangkan, maka
diperlukan begitu banyak penelitian dari banyak aspek,
salah satunya adalah mengidentifikasi miRNA yang
berkaitan dengan proses perkembangan virus. Pada
penelitian ini didapatkan lima miRNA yang sangat berperan
pada infeksi virus SARS-CoV-2 melalui analisis
bioinformatik sehingga sangat diperlukan penelitian lebih
lanjut untuk melihat kelima miRNA tersebut pada sampel
manusia yang terinfeksi SARS-CoV-2.
Rencana Tindak
Lanjut
Penelitian ini perlu ditindaklanjuti pada sampel darah
manusia yang terinfeksi virus SARS-CoV-2 untuk melihat
apakah miRNA yang kami temukan dalam penelitian ini
mengalami peningkatan atau tidak, sehingga dapat
dimanfaatkan untuk biomarker potensial atau untuk tujuan
pengobatan pada kasus COVID-19 di Indonesia.

21
DAFTAR PUSTAKA
Abu-Farha, M., Thanaraj, T. A., Qaddoumi, M. G., Hashem, A., Abubaker, J., &
Al-Mulla, F. (2020). The Role of Lipid Metabolism in COVID-19 Virus
Infection and as a Drug Target. International Journal of Molecular Sciences,
21(10), 3544. https://doi.org/10.3390/ijms21103544
Alm, E., Broberg, E. K., Connor, T., Hodcroft, E. B., Komissarov, A. B., Maurer-
Stroh, S., Melidou, A., Neher, R. A., O’Toole, Á., & Pereyaslov, D. (2020).
Geographical and temporal distribution of SARS-CoV-2 clades in the WHO
European Region, January to June 2020. Eurosurveillance, 25(32).
https://doi.org/10.2807/1560-7917.ES.2020.25.32.2001410
Ansori, A. N., Kusala, M., Irine, N., Indrasari, S., Alamudi, Muhammad Y Nidom,
R. V, & Al., E. (2020). Immunoinformatic investigation of three structural
protein genes in Indonesia SARS-CoV-2 isolates. Systematic Reviews in
Pharmacy, 11(7), 422–434.
Arisan, E. D., Dart, A., Grant, G. H., Arisan, S., Cuhadaroglu, S., Lange, S., &
Uysal-Onganer, P. (2020). The Prediction of miRNAs in SARS-CoV-2
Genomes: hsa-miR Databases Identify 7 Key miRs Linked to Host Responses
and Virus Pathogenicity-Related KEGG Pathways Significant for
Comorbidities. Viruses, 12(6), 614. https://doi.org/10.3390/v12060614
Bickels, J., Jelinek, J., Shmookler, B., & Malawer, M. (2001). Biopsy of
Musculoskeletal Tumors. In Musculoskeletal Cancer Surgery (pp. 37–46).
Bouhaddou, M., Memon, D., Meyer, B., White, K. M., Rezelj, V. V., Correa
Marrero, M., Polacco, B. J., Melnyk, J. E., Ulferts, S., Kaake, R. M., Batra, J.,
Richards, A. L., Stevenson, E., Gordon, D. E., Rojc, A., Obernier, K., Fabius,
J. M., Soucheray, M., Miorin, L., … Krogan, N. J. (2020). The Global
Phosphorylation Landscape of SARS-CoV-2 Infection. Cell, 182(3), 685-
712.e19. https://doi.org/10.1016/j.cell.2020.06.034
Cammaerts, S., Strazisar, M., De Rijk, P., & Del Favero, J. (2015). Genetic variants
in microRNA genes: impact on microRNA expression, function, and disease.
Frontiers in Genetics, 6(186). https://doi.org/10.3389/fgene.2015.00186.
Cao, Y., Li, L., Feng, Z., Wan, S., Huang, P., Sun, X., Wen, F., Huang, X., Ning,
G., & Wang, W. (2020). Comparative genetic analysis of the novel
coronavirus (2019-nCoV/SARS-CoV-2) receptor ACE2 in different
populations. Cell Discovery, 6(1), 11. https://doi.org/10.1038/s41421-020-
0147-1
Chan, J. F.-W., Yuan, S., Kok, K.-H., To, K. K.-W., Chu, H., Yang, J., Xing, F.,
Liu, J., Yip, C. C.-Y., Poon, R. W.-S., Tsoi, H.-W., Lo, S. K.-F., Chan, K.-H.,
Poon, V. K.-M., Chan, W.-M., Ip, J. D., Cai, J.-P., Cheng, V. C.-C., Chen, H.,
… Yuen, K.-Y. (2020). A familial cluster of pneumonia associated with the
2019 novel coronavirus indicating person-to-person transmission: a study of a

22
family cluster. The Lancet, 395(10223), 514–523.
https://doi.org/10.1016/S0140-6736(20)30154-9
Chen, N., Zhou, M., Dong, X., Qu, J., Gong, F., Han, Y., Qiu, Y., Wang, J., Liu,
Y., Wei, Y., Xia, J., Yu, T., Zhang, X., & Zhang, L. (2020). Epidemiological
and clinical characteristics of 99 cases of 2019 novel coronavirus pneumonia
in Wuhan, China: a descriptive study. The Lancet, 395(10223), 507–513.
https://doi.org/10.1016/S0140-6736(20)30211-7
Cummins, J. M., He, Y., Leary, R. J., Pagliarini, R., Diaz, L. A., Sjoblom, T., Barad,
O., Bentwich, Z., Szafranska, A. E., Labourier, E., Raymond, C. K., Roberts,
B. S., Juhl, H., Kinzler, K. W., Vogelstein, B., & Velculescu, V. E. (2006).
The colorectal microRNAome. Proceedings of the National Academy of
Sciences, 103(10), 3687–3692. https://doi.org/10.1073/pnas.0511155103
Das, U. N. (2020). Can Bioactive Lipids Inactivate Coronavirus (COVID-19)?
Archives of Medical Research, 51(3), 282–286.
https://doi.org/10.1016/j.arcmed.2020.03.004
de Wit, E., van Doremalen, N., Falzarano, D., & Munster, V. J. (2016). SARS and
MERS: recent insights into emerging coronaviruses. Nature Reviews
Microbiology, 14(8), 523–534. https://doi.org/10.1038/nrmicro.2016.81
Demirci, M. D. S., Toprak, M., & Allmer, J. (2016). A Machine Learning Approach
for MicroRNA Precursor Prediction in Retro-transcribing Virus Genomes.
Journal of Integrative Bioinformatics, 13(5). https://doi.org/10.1515/jib-2016-
303
Fernandez, N., Cordiner, R. A., Young, R. S., Hug, N., Macias, S., & Cáceres, J. F.
(2017). Genetic variation and RNA structure regulate microRNA biogenesis.
Nature Communications, 8(1), 15114. https://doi.org/10.1038/ncomms15114
Finnegan, E. F., & Pasquinelli, A. E. (2013). MicroRNA biogenesis: regulating the
regulators. Critical Reviews in Biochemistry and Molecular Biology, 48(1),
51–68. https://doi.org/10.3109/10409238.2012.738643
Franco-Muñoz, C., Álvarez-Díaz, D. A., Laiton-Donato, K., Wiesner, M.,
Escandón, P., Usme-Ciro, J. A., Franco-Sierra, N. D., Flórez-Sánchez, A. C.,
Gómez-Rangel, S., Rodríguez-Calderon, L. D., Barbosa-Ramirez, J., Ospitia-
Baez, E., Walteros, D. M., Ospina-Martinez, M. L., & Mercado-Reyes, M.
(2020). Substitutions in Spike and Nucleocapsid proteins of SARS-CoV-2
circulating in South America. Infection, Genetics and Evolution, 85, 104557.
https://doi.org/10.1016/j.meegid.2020.104557
Fulzele, S., Sahay, B., Yusufu, I., Lee, T. J., Sharma, A., Kolhe, R., & Isales, C. M.
(2020). COVID-19 Virulence in Aged Patients Might Be Impacted by the Host
Cellular MicroRNAs Abundance/Profile. Aging and Disease, 11(3), 509.
https://doi.org/10.14336/AD.2020.0428
Geurdes, H., & Koutsaroff, I. (2020). Histamine Antagonists to Temper the

23
Cytokine Overproduction in Gastrointestinal Cells Infected by SARS-CoV-2.
Virol Mycol Open Access, 9(2), 189.
https://doi.org/https://doi.org/10.20944/preprints202004.0542.v1
Gkirtzou, K., Tsamardinos, I., Tsakalides, P., & Poirazi, P. (2010). MatureBayes:
A Probabilistic Algorithm for Identifying the Mature miRNA within Novel
Precursors. PLoS ONE, 5(8), e11843.
https://doi.org/10.1371/journal.pone.0011843
Grundhoff, A. (2011). Computational Prediction of Viral miRNAs (pp. 143–152).
https://doi.org/10.1007/978-1-61779-037-9_8
Guan, W., Ni, Z., Hu, Y., Liang, W., Ou, C., He, J., Liu, L., Shan, H., Lei, C., Hui,
D. S. C., Du, B., Li, L., Zeng, G., Yuen, K.-Y., Chen, R., Tang, C., Wang, T.,
Chen, P., Xiang, J., … Zhong, N. (2020). Clinical Characteristics of
Coronavirus Disease 2019 in China. New England Journal of Medicine,
382(18), 1708–1720. https://doi.org/10.1056/NEJMoa2002032
gugus tugas percepatan penanganan covid 19. (n.d.). https://covid19.go.id/
Haddad, H., & Walid Al-Zyoud. (2020). miRNA target prediction might explain
the reduced transmission of SARS-CoV-2 in Jordan, Middle East. Non-Coding
RNA Research, 5(3), 135–143. https://doi.org/10.1016/j.ncrna.2020.08.002
Hamed S, Elkhatib W, Khairallah A, N. A. (2020). Global dynamics of SARS-CoV-
2 clades and their relation to COVID-19 epidemiology 2020. Research Square.
https://doi.org/https://doi.org/10.21203/rs.3.rs-89876/v1
Hartley, P. D., Tillett, R. L., AuCoin, D. P., Sevinsky, J. R., Xu, Y., Gorzalski, A.,
Pandori, M., Buttery, E., Hansen, H., Picker, M. A., Rossetto, C. C., & Verma,
S. C. (2020). Genomic surveillance of Nevada patients revealed prevalence of
unique SARS-CoV-2 variants bearing mutations in the RdRp gene. MedRxiv.
https://doi.org/https://doi.org/10.1101/2020.08.21.20178863
Hassan, S. S., Choudhury, P. P., Roy, B., & Jana, S. S. (2020). Missense mutations
in SARS-CoV2 genomes from Indian patients. Genomics, 112(6), 4622–4627.
https://doi.org/10.1016/j.ygeno.2020.08.021
Hoffmann, M., Kleine-Weber, H., Schroeder, S., Krüger, N., Herrler, T., Erichsen,
S., Schiergens, T. S., Herrler, G., Wu, N.-H., Nitsche, A., Müller, M. A.,
Drosten, C., & Pöhlmann, S. (2020). SARS-CoV-2 Cell Entry Depends on
ACE2 and TMPRSS2 and Is Blocked by a Clinically Proven Protease
Inhibitor. Cell, 181(2), 271-280.e8. https://doi.org/10.1016/j.cell.2020.02.052
Hoxha, M. (2020). What about COVID-19 and arachidonic acid pathway?
European Journal of Clinical Pharmacology, 76(11), 1501–1504.
https://doi.org/10.1007/s00228-020-02941-w
Hu, B., Guo, H., Zhou, P., & Shi, Z.-L. (2020). Characteristics of SARS-CoV-2 and
COVID-19. Nature Reviews Microbiology. https://doi.org/10.1038/s41579-
020-00459-7

24
Hu, X., Cai, X., Song, X., Li, C., Zhao, J., Luo, W., Zhang, Q., Ekumi, I. O., & He,
Z. (2020). Possible SARS-coronavirus 2 inhibitor revealed by simulated
molecular docking to viral main protease and host toll-like receptor. Future
Virology, 15(6), 359–368. https://doi.org/10.2217/fvl-2020-0099
Huang, C., Wang, Y., Li, X., Ren, L., Zhao, J., Hu, Y., Zhang, L., Fan, G., Xu, J.,
Gu, X., Cheng, Z., Yu, T., Xia, J., Wei, Y., Wu, W., Xie, X., Yin, W., Li, H.,
Liu, M., … Cao, B. (2020). Clinical features of patients infected with 2019
novel coronavirus in Wuhan, China. The Lancet, 395(10223), 497–506.
https://doi.org/10.1016/S0140-6736(20)30183-5
Jackson, C. B., Zhang, L., Farzan, M., & Choe, H. (2020). Functional importance
of the D614G mutation in the SARS-CoV-2 spike protein. Biochemical and
Biophysical Research Communications.
https://doi.org/10.1016/j.bbrc.2020.11.026
Jima, D. D., Zhang, J., Jacobs, C., Richards, K. L., Dunphy, C. H., Choi, W. W. L.,
Yan Au, W., Srivastava, G., Czader, M. B., Rizzieri, D. A., Lagoo, A. S.,
Lugar, P. L., Mann, K. P., Flowers, C. R., Bernal-Mizrachi, L., Naresh, K. N.,
Evens, A. M., Gordon, L. I., Luftig, M., … Dave, S. S. (2010). Deep
sequencing of the small RNA transcriptome of normal and malignant human
B cells identifies hundreds of novel microRNAs. Blood, 116(23), e118–e127.
https://doi.org/10.1182/blood-2010-05-285403
Kannan, S. R., Spratt, A. N., Quinn, T. P., Heng, X., Lorson, C. L., Sönnerborg, A.,
Byrareddy, S. N., & Singh, K. (2020). Infectivity of SARS-CoV-2: there Is
Something More than D614G? Journal of Neuroimmune Pharmacology,
15(4), 574–577. https://doi.org/10.1007/s11481-020-09954-3
Khan, M. A.-A.-K., Sany, M. R. U., Islam, M. S., & Islam, A. B. M. M. K. (2020).
Epigenetic Regulator miRNA Pattern Differences Among SARS-CoV, SARS-
CoV-2, and SARS-CoV-2 World-Wide Isolates Delineated the Mystery
Behind the Epic Pathogenicity and Distinct Clinical Characteristics of
Pandemic COVID-19. Frontiers in Genetics, 11.
https://doi.org/10.3389/fgene.2020.00765
Korber, B., Fischer, W. M., Gnanakaran, S., Yoon, H., Theiler, J., Abfalterer, W.,
Hengartner, N., Giorgi, E. E., Bhattacharya, T., Foley, B., Hastie, K. M.,
Parker, M. D., Partridge, D. G., Evans, C. M., Freeman, T. M., de Silva, T. I.,
McDanal, C., Perez, L. G., Tang, H., … Wyles, M. D. (2020). Tracking
Changes in SARS-CoV-2 Spike: Evidence that D614G Increases Infectivity
of the COVID-19 Virus. Cell, 182(4), 812-827.e19.
https://doi.org/10.1016/j.cell.2020.06.043
Kothapalli, K. S. D., Park, H. G., & Brenna, J. T. (2020). Polyunsaturated fatty acid
biosynthesis pathway and genetics. implications for interindividual variability
in prothrombotic, inflammatory conditions such as COVID-19.
Prostaglandins, Leukotrienes and Essential Fatty Acids, 162, 102183.
https://doi.org/10.1016/j.plefa.2020.102183

25
Koyama, T., Platt, D., & Parida, L. (2020). Variant analysis of SARS-CoV-2
genomes. Bulletin of the World Health Organization, 98(7), 495–504.
https://doi.org/10.2471/BLT.20.253591
Krol, J., Loedige, I., & Filipowicz, W. (2010). The widespread regulation of
microRNA biogenesis, function and decay. Nature Reviews Genetics, 11(9),
597–610. https://doi.org/10.1038/nrg2843
Kuba, K., Imai, Y., Rao, S., Gao, H., Guo, F., Guan, B., Huan, Y., Yang, P., Zhang,
Y., Deng, W., Bao, L., Zhang, B., Liu, G., Wang, Z., Chappell, M., Liu, Y.,
Zheng, D., Leibbrandt, A., Wada, T., … Penninger, J. M. (2005). A crucial
role of angiotensin converting enzyme 2 (ACE2) in SARS coronavirus–
induced lung injury. Nature Medicine, 11(8), 875–879.
https://doi.org/10.1038/nm1267
Leng, L., Cao, R., Ma, J., Mou, D., Zhu, Y., Li, W., Lv, L., Gao, D., Zhang, S.,
Gong, F., Zhao, L., Qiu, B., Xiang, H., Hu, Z., Feng, Y., Dai, Y., Zhao, J., Wu,
Z., Li, H., & Zhong, W. (2020). Pathological features of COVID-19-associated
lung injury: a preliminary proteomics report based on clinical samples. Signal
Transduction and Targeted Therapy, 5(1), 240.
https://doi.org/10.1038/s41392-020-00355-9
Li, C., Hu, X., Li, L., & Li, J. (2020). Differential microRNA expression in the
peripheral blood from human patients with COVID‐19. Journal of Clinical
Laboratory Analysis, 34(10). https://doi.org/10.1002/jcla.23590
Liu, Z., Wang, J., Xu, Y., Guo, M., Mi, K., Xu, R., Pei, Y., Zhang, Q., Luan, X.,
Hu, Z., & Liu1, X. (2020). Implications of the virus-encoded miRNA and host
miRNA in the pathogenicity of SARS-CoV-2.
Martin-Guerrero, I., Gutierrez-Camino, A., Lopez-Lopez, E., Bilbao-Aldaiturriaga,
N., Pombar-Gomez, M., Ardanaz, M., & Garcia-Orad, A. (2015). Genetic
Variants in MiRNA Processing Genes and Pre-MiRNAs Are Associated with
the Risk of Chronic Lymphocytic Leukemia. PLOS ONE, 10(3), e0118905.
https://doi.org/10.1371/journal.pone.0118905
Mendell, J. T., & Olson, E. N. (2012). MicroRNAs in Stress Signaling and Human
Disease. Cell, 148(6), 1172–1187. https://doi.org/10.1016/j.cell.2012.02.005
Mercatelli, D., & Giorgi, F. M. (2020). Geographic and Genomic Distribution of
SARS-CoV-2 Mutations. Frontiers in Microbiology, 11.
https://doi.org/10.3389/fmicb.2020.01800
Mohammad, A., Alshawaf, E., Marafie, S. K., Abu-Farha, M., Abubaker, J., & Al-
Mulla, F. (2020). Higher binding affinity of Furin to SARS-CoV-2 spike (S)
protein D614G could be associated with higher SARS-CoV-2 infectivity.
International Journal of Infectious Diseases.
https://doi.org/10.1016/j.ijid.2020.10.033
Muhammad Ansori, A. N., Dhea Kharisma, V., Sabilil Muttaqin, S., Antonius, Y.,

26
& Parikesit, A. A. (2020). Genetic Variant of SARS-CoV-2 Isolates in
Indonesia: Spike Glycoprotein Gene. Journal of Pure and Applied
Microbiology, 14(suppl 1), 971–978.
https://doi.org/10.22207/JPAM.14.SPL1.35
Mutlu, O., Ugurel, O. M., Sariyer, E., Ata, O., Inci, T. G., Ugurel, E., Kocer, S., &
Turgut-Balik, D. (2020). Targeting SARS-CoV-2 Nsp12/Nsp8 interaction
interface with approved and investigational drugs: an in silico structure-based
approach. Journal of Biomolecular Structure and Dynamics, 1–13.
https://doi.org/10.1080/07391102.2020.1819882
Nguyen, T. T., Pham, T. N., Van, T. D., Nguyen, T. T., Nguyen, D. T. N., Le, H.
N. M., Eden, J.-S., Rockett, R. J., Nguyen, T. T. H., Vu, B. T. N., Tran, G.
Van, Le, T. Van, Dwyer, D. E., & van Doorn, H. R. (2020). Genetic diversity
of SARS-CoV-2 and clinical, epidemiological characteristics of COVID-19
patients in Hanoi, Vietnam. PLOS ONE, 15(11), e0242537.
https://doi.org/10.1371/journal.pone.0242537
Nidom R V, Indrasari S, Normalina I, Kusala MKJ, Ansori ANM, N. C. (2020).
Investigation of the D614G Mutation and Antibody-Dependent Enhancement
Sequences in Indonesia SARSCoV-2 Isolates and Comparison to Southeast
Asian Isolates. Systematic Reviews in Pharmacy, 11(8), 203–213.
https://doi.org/http://dx.doi.org/10.31838/srp.2020.8.30
Ouyang, Y., Yin, J., Wang, W., Shi, H., Shi, Y., Xu, B., Qiao, L., Feng, Y., Pang,
L., Wei, F., Guo, X., Jin, R., & Chen, D. (2020). Downregulated Gene
Expression Spectrum and Immune Responses Changed During the Disease
Progression in Patients With COVID-19. Clinical Infectious Diseases, 71(16),
2052–2060. https://doi.org/10.1093/cid/ciaa462
Paolini, R., Numerof, R., & Kinet, J. P. (1992). Phosphorylation/dephosphorylation
of high-affinity IgE receptors: a mechanism for coupling/uncoupling a large
signaling complex. Proceedings of the National Academy of Sciences, 89(22),
10733–10737. https://doi.org/10.1073/pnas.89.22.10733
Paraskevopoulou, M. D., Georgakilas, G., Kostoulas, N., Vlachos, I. S., Vergoulis,
T., Reczko, M., Filippidis, C., Dalamagas, T., & Hatzigeorgiou, A. G. (2013).
DIANA-microT web server v5.0: service integration into miRNA functional
analysis workflows. Nucleic Acids Research, 41(W1), W169–W173.
https://doi.org/10.1093/nar/gkt393
Parikesit, Arli Aditya, & Nurdiansyah, R. (2020). The predicted structure for the
anti-sense sirna of the RNA Polymerase Enzym (RDRP) gene of the SAR-
COV-2. Berita Biologi, 19(1), 97–108.
https://doi.org/10.14203/beritabiologi.v19i1.3849
Parikesit, Arli Ardi, & Nurdiansyah, R. (2020). Drug Repurposing Option for
COVID-19 with Structural Bioinformatics of Chemical Interactions
Approach. Cermin Dunia Kedokteran, 47(3), 222–226.

27
Pedoman pencegahan dan pengendalian corona virus disease (COVID - 19).
(2020). Kementerian Kesehatan RI Direktorat Jenderal Pencegahan dan
Pengendalian Penyakit (P2P).
Persson, H., Kvist, A., Rego, N., Staaf, J., Vallon-Christersson, J., Luts, L., Loman,
N., Jonsson, G., Naya, H., Hoglund, M., Borg, A., & Rovira, C. (2011).
Identification of New MicroRNAs in Paired Normal and Tumor Breast Tissue
Suggests a Dual Role for the ERBB2/Her2 Gene. Cancer Research, 71(1), 78–
86. https://doi.org/10.1158/0008-5472.CAN-10-1869
Pfeffer, S. (2004). Identification of Virus-Encoded MicroRNAs. Science,
304(5671), 734–736. https://doi.org/10.1126/science.1096781
Plante, J. A., Liu, Y., Liu, J., Xia, H., Johnson, B. A., Lokugamage, K. G., Zhang,
X., Muruato, A. E., Zou, J., Fontes-Garfias, C. R., Mirchandani, D., Scharton,
D., Bilello, J. P., Ku, Z., An, Z., Kalveram, B., Freiberg, A. N., Menachery, V.
D., Xie, X., … Shi, P.-Y. (2020). Spike mutation D614G alters SARS-CoV-2
fitness and neutralization susceptibility. BioRxiv.
https://doi.org/https://doi.org/10.1101/2020.09.01.278689
Pyrc, K., Dijkman, R., Deng, L., Jebbink, M. F., Ross, H. A., Berkhout, B., & van
der Hoek, L. (2006). Mosaic Structure of Human Coronavirus NL63, One
Thousand Years of Evolution. Journal of Molecular Biology, 364(5), 964–973.
https://doi.org/10.1016/j.jmb.2006.09.074
Saçar Demirci, M. D., & Adan, A. (2020). Computational analysis of microRNA-
mediated interactions in SARS-CoV-2 infection. PeerJ, 8, e9369.
https://doi.org/10.7717/peerj.9369
Sarma, A., Phukan, H., Halder, N., & Madanan, M. G. (2020). An in-silico approach
to study the possible interactions of miRNA between human and SARS-CoV2.
Computational Biology and Chemistry, 88, 107352.
https://doi.org/10.1016/j.compbiolchem.2020.107352
Shawky, E., Nada, A. A., & Ibrahim, R. S. (2020). Potential role of medicinal plants
and their constituents in the mitigation of SARS-CoV-2: identifying related
therapeutic targets using network pharmacology and molecular docking
analyses. RSC Advances, 10(47), 27961–27983.
https://doi.org/10.1039/D0RA05126H
Siraganian, R. (2003). Mast cell signal transduction from the high-affinity IgE
receptor. Current Opinion in Immunology, 15(6), 639–646.
https://doi.org/10.1016/j.coi.2003.09.010
Skalsky, R. L., & Cullen, B. R. (2010). Viruses, microRNAs, and Host Interactions.
Annual Review of Microbiology, 64(1), 123–141.
https://doi.org/10.1146/annurev.micro.112408.134243
Soratto, T. A. T., Darban, H., Bjerkner, A., Coorens, M., Albert, J., Allander, T., &
Andersson, B. (2020). Four SARS-CoV-2 Genome Sequences from Late April

28
in Stockholm, Sweden, Reveal a Rare Mutation in the Spike Protein.
Microbiology Resource Announcements, 9(35).
https://doi.org/10.1128/MRA.00934-20
Sun, Z., Ren, K., Zhang, X., Chen, J., Jiang, Z., Jiang, J., Ji, F., Ouyang, X., & Li,
L. (2020). Mass Spectrometry Analysis of Newly Emerging Coronavirus
HCoV-19 Spike Protein and Human ACE2 Reveals Camouflaging Glycans
and Unique Post-Translational Modifications. Engineering.
https://doi.org/10.1016/j.eng.2020.07.014
Thomas, T., Stefanoni, D., Reisz, J. A., Nemkov, T., Bertolone, L., Francis, R. O.,
Hudson, K. E., Zimring, J. C., Hansen, K. C., Hod, E. A., Spitalnik, S. L., &
D’Alessandro, A. (2020). COVID-19 infection alters kynurenine and fatty acid
metabolism, correlating with IL-6 levels and renal status. JCI Insight, 5(14).
https://doi.org/10.1172/jci.insight.140327
Trobaugh, D. W., & Klimstra, W. B. (2017). MicroRNA Regulation of RNA Virus
Replication and Pathogenesis. Trends in Molecular Medicine, 23(1), 80–93.
https://doi.org/10.1016/j.molmed.2016.11.003
Tu, Y.-F., Chien, C.-S., Yarmishyn, A. A., Lin, Y.-Y., Luo, Y.-H., Lin, Y.-T., Lai,
W.-Y., Yang, D.-M., Chou, S.-J., Yang, Y.-P., Wang, M.-L., & Chiou, S.-H.
(2020). A Review of SARS-CoV-2 and the Ongoing Clinical Trials.
International Journal of Molecular Sciences, 21(7), 2657.
https://doi.org/10.3390/ijms21072657
Valko, M., Leibfritz, D., Moncol, J., Cronin, M. T. D., Mazur, M., & Telser, J.
(2007). Free radicals and antioxidants in normal physiological functions and
human disease. The International Journal of Biochemistry & Cell Biology,
39(1), 44–84. https://doi.org/10.1016/j.biocel.2006.07.001
Vaz, C., Ahmad, H. M., Sharma, P., Gupta, R., Kumar, L., Kulshreshtha, R., &
Bhattacharya, A. (2010). Analysis of microRNA transcriptome by deep
sequencing of small RNA libraries of peripheral blood. BMC Genomics, 11(1),
288. https://doi.org/10.1186/1471-2164-11-288
Vlachos, I. S., Zagganas, K., Paraskevopoulou, M. D., Georgakilas, G.,
Karagkouni, D., Vergoulis, T., Dalamagas, T., & Hatzigeorgiou, A. G. (2015).
DIANA-miRPath v3.0: deciphering microRNA function with experimental
support. Nucleic Acids Research, 43(W1), W460–W466.
https://doi.org/10.1093/nar/gkv403
Wang, R., Chen, J., Gao, K., Hozumi, Y., Yin, C., & Wei, G.-W. (2020).
Characterizing SARS-CoV-2 mutations in the United States. ArXiv.
https://doi.org/https://www.x-
mol.com/paperRedirect/1304348658974822400
Wang, X. (2008). miRDB: A microRNA target prediction and functional annotation
database with a wiki interface. RNA, 14(6), 1012–1017.
https://doi.org/10.1261/rna.965408

29
WHO. (n.d.). Coronavirus disease 2019 (COVID-19) 2020.
https://www.who.int/indonesia/news/novel-coronavirus/situation-reports
Wimmer, E., & Goldbach, R. (1992). Viral genetics. Current Opinion in Genetics
& Development, 2(1), 59–60. https://doi.org/10.1016/S0959-437X(05)80322-
3
Winter, J., Jung, S., Keller, S., Gregory, R. I., & Diederichs, S. (2009). Many roads
to maturity: microRNA biogenesis pathways and their regulation. Nature Cell
Biology, 11(3), 228–234. https://doi.org/10.1038/ncb0309-228
Wu, F., Zhao, S., Yu, B., Chen, Y.-M., Wang, W., Song, Z.-G., Hu, Y., Tao, Z.-W.,
Tian, J.-H., Pei, Y.-Y., Yuan, M.-L., Zhang, Y.-L., Dai, F.-H., Liu, Y., Wang,
Q.-M., Zheng, J.-J., Xu, L., Holmes, E. C., & Zhang, Y.-Z. (2020). A new
coronavirus associated with human respiratory disease in China. Nature,
579(7798), 265–269. https://doi.org/10.1038/s41586-020-2008-3
Xia HP. (2008). Great potential of microRNA in cancer stem cell. J Cancer Mol,
4(3), 79–89.
Yurkovetskiy, L., Wang, X., Pascal, K. E., Tomkins-Tinch, C., Nyalile, T. P.,
Wang, Y., Baum, A., Diehl, W. E., Dauphin, A., Carbone, C., Veinotte, K.,
Egri, S. B., Schaffner, S. F., Lemieux, J. E., Munro, J. B., Rafique, A., Barve,
A., Sabeti, P. C., Kyratsous, C. A., … Luban, J. (2020). Structural and
Functional Analysis of the D614G SARS-CoV-2 Spike Protein Variant. Cell,
183(3), 739-751.e8. https://doi.org/10.1016/j.cell.2020.09.032

30
LAMPIRAN
31

32
33
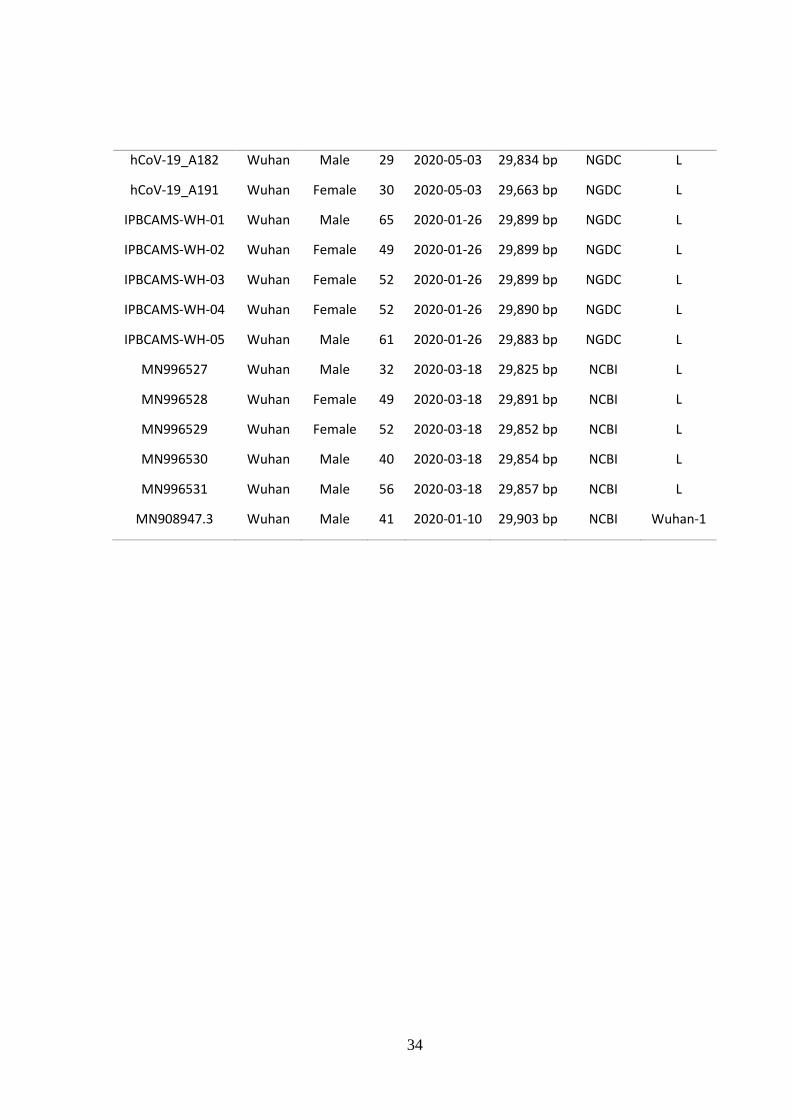
Tabel data tambahan 1. Sequence SARS-CoV-2 dari sampel Wuhan
Sequence_ID Location Gender Age Submission
Date
Sequence
Length
Database Clade
EPI_ISL_402120 Wuhan Male 61 2020-01-10 29,896 bp GSAID L
EPI_ISL_402121 Wuhan Male 32 2020-01-11 29,891 bp GSAID L
EPI_ISL_402132 Wuhan Female 49 2020-01-19 29,848 bp GSAID L
EPI_ISL_406798 Wuhan Male 44 2020-01-30 29,866 bp GSAID L
EPI_ISL_406800 Wuhan Female 21 2020-01-30 29,868 bp GSAID L
EPI_ISL_406801 Wuhan Male 39 2020-01-30 29,872 bp GSAID S
EPI_ISL_412899 Wuhan Female 49 2020-02-28 29,848 bp GSAID L
EPI_ISL_434534 Wuhan Male 32 2020-04-28 29,892 bp GSAID L
EPI_ISL_493166 Wuhan Female 54 2020-07-21 29,663 bp GSAID L
hCoV-19_A137 Wuhan Female 47 2020-05-03 29,663 bp NGDC L
hCoV-19_A142 Wuhan Female 48 2020-05-03 29,663 bp NGDC L
hCoV-19_A145 Wuhan Female 41 2020-05-03 29,663 bp NGDC L
hCoV-19_A148 Wuhan Male 44 2020-05-03 29,663 bp NGDC L
hCoV-19_A152 Wuhan Male 53 2020-05-03 29,663 bp NGDC L
hCoV-19_A153 Wuhan Female 29 2020-05-03 29,663 bp NGDC L
hCoV-19_A160 Wuhan Female 30 2020-05-03 29,835 bp NGDC L
hCoV-19_A163 Wuhan Female 48 2020-05-03 29,834 bp NGDC L
hCoV-19_A164 Wuhan Female 52 2020-05-03 29,834 bp NGDC L
hCoV-19_A167 Wuhan Male 23 2020-05-03 29,834 bp NGDC L
hCoV-19_A168 Wuhan Female 47 2020-05-03 29,834 bp NGDC L
hCoV-19_A169 Wuhan Female 25 2020-05-03 29,836 bp NGDC L
hCoV-19_A170 Wuhan Female 56 2020-05-03 29,834 bp NGDC L
hCoV-19_A171 Wuhan Male 43 2020-05-03 29,833 bp NGDC L
hCoV-19_A174 Wuhan Male 60 2020-05-03 29,663 bp NGDC L
34
hCoV-19_A182 Wuhan Male 29 2020-05-03 29,834 bp NGDC L
hCoV-19_A191 Wuhan Female 30 2020-05-03 29,663 bp NGDC L
IPBCAMS-WH-01 Wuhan Male 65 2020-01-26 29,899 bp NGDC L
IPBCAMS-WH-02 Wuhan Female 49 2020-01-26 29,899 bp NGDC L
IPBCAMS-WH-03 Wuhan Female 52 2020-01-26 29,899 bp NGDC L
IPBCAMS-WH-04 Wuhan Female 52 2020-01-26 29,890 bp NGDC L
IPBCAMS-WH-05 Wuhan Male 61 2020-01-26 29,883 bp NGDC L
MN996527 Wuhan Male 32 2020-03-18 29,825 bp NCBI L
MN996528 Wuhan Female 49 2020-03-18 29,891 bp NCBI L
MN996529 Wuhan Female 52 2020-03-18 29,852 bp NCBI L
MN996530 Wuhan Male 40 2020-03-18 29,854 bp NCBI L
MN996531 Wuhan Male 56 2020-03-18 29,857 bp NCBI L
MN908947.3 Wuhan Male 41 2020-01-10 29,903 bp NCBI Wuhan-1
35
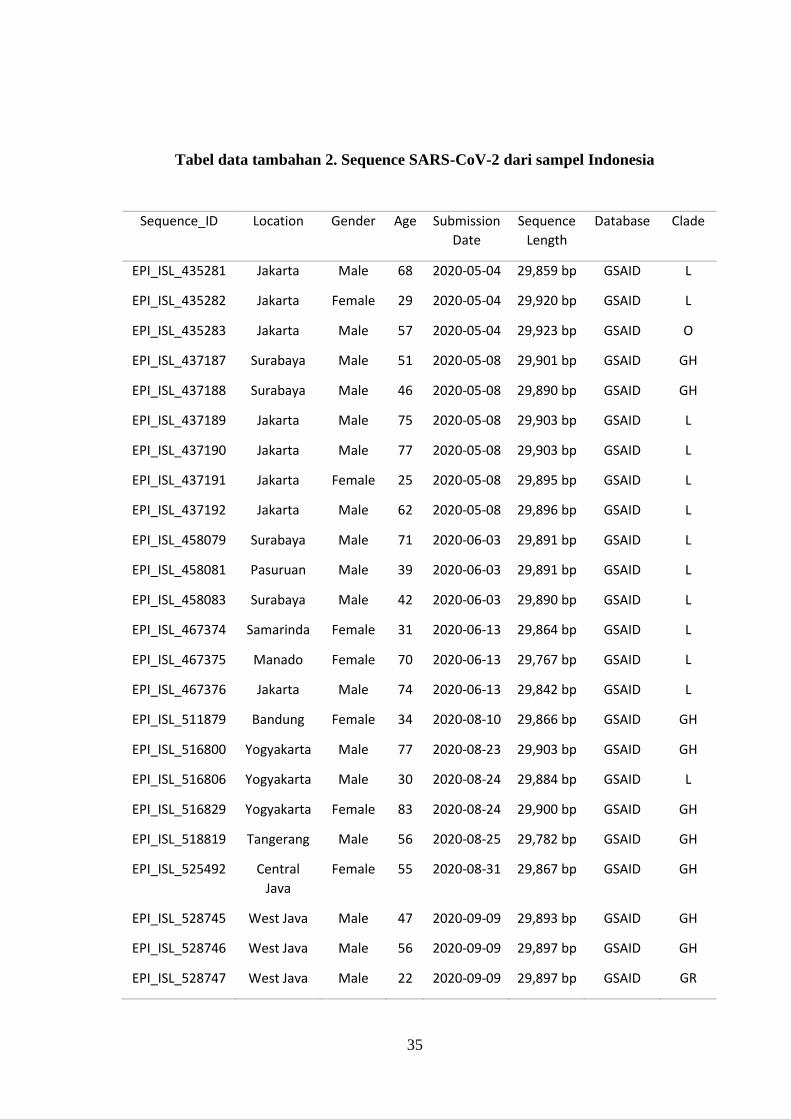
Tabel data tambahan 2. Sequence SARS-CoV-2 dari sampel Indonesia
Sequence_ID Location Gender Age Submission
Date
Sequence
Length
Database Clade
EPI_ISL_435281 Jakarta Male 68 2020-05-04 29,859 bp GSAID L
EPI_ISL_435282 Jakarta Female 29 2020-05-04 29,920 bp GSAID L
EPI_ISL_435283 Jakarta Male 57 2020-05-04 29,923 bp GSAID O
EPI_ISL_437187 Surabaya Male 51 2020-05-08 29,901 bp GSAID GH
EPI_ISL_437188 Surabaya Male 46 2020-05-08 29,890 bp GSAID GH
EPI_ISL_437189 Jakarta Male 75 2020-05-08 29,903 bp GSAID L
EPI_ISL_437190 Jakarta Male 77 2020-05-08 29,903 bp GSAID L
EPI_ISL_437191 Jakarta Female 25 2020-05-08 29,895 bp GSAID L
EPI_ISL_437192 Jakarta Male 62 2020-05-08 29,896 bp GSAID L
EPI_ISL_458079 Surabaya Male 71 2020-06-03 29,891 bp GSAID L
EPI_ISL_458081 Pasuruan Male 39 2020-06-03 29,891 bp GSAID L
EPI_ISL_458083 Surabaya Male 42 2020-06-03 29,890 bp GSAID L
EPI_ISL_467374 Samarinda Female 31 2020-06-13 29,864 bp GSAID L
EPI_ISL_467375 Manado Female 70 2020-06-13 29,767 bp GSAID L
EPI_ISL_467376 Jakarta Male 74 2020-06-13 29,842 bp GSAID L
EPI_ISL_511879 Bandung Female 34 2020-08-10 29,866 bp GSAID GH
EPI_ISL_516800 Yogyakarta Male 77 2020-08-23 29,903 bp GSAID GH
EPI_ISL_516806 Yogyakarta Male 30 2020-08-24 29,884 bp GSAID L
EPI_ISL_516829 Yogyakarta Female 83 2020-08-24 29,900 bp GSAID GH
EPI_ISL_518819 Tangerang Male 56 2020-08-25 29,782 bp GSAID GH
EPI_ISL_525492 Central
Java
Female 55 2020-08-31 29,867 bp GSAID GH
EPI_ISL_528745 West Java Male 47 2020-09-09 29,893 bp GSAID GH
EPI_ISL_528746 West Java Male 56 2020-09-09 29,897 bp GSAID GH
EPI_ISL_528747 West Java Male 22 2020-09-09 29,897 bp GSAID GR
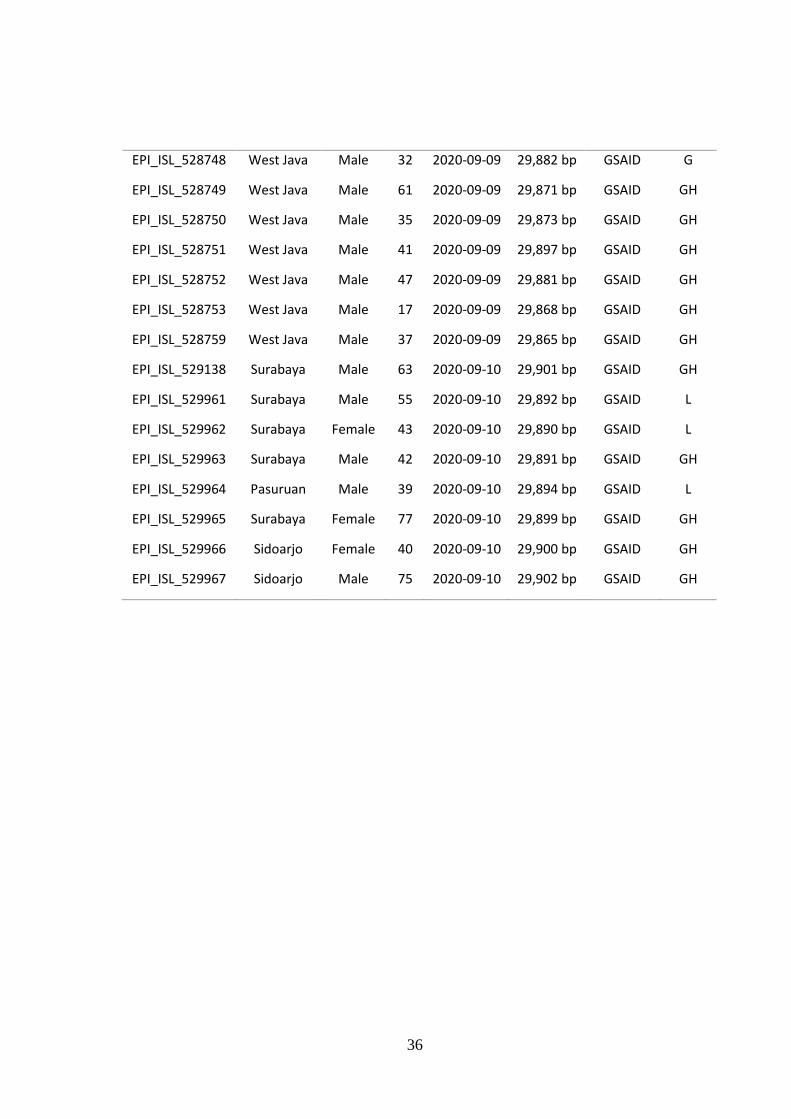
36
EPI_ISL_528748 West Java Male 32 2020-09-09 29,882 bp GSAID G
EPI_ISL_528749 West Java Male 61 2020-09-09 29,871 bp GSAID GH
EPI_ISL_528750 West Java Male 35 2020-09-09 29,873 bp GSAID GH
EPI_ISL_528751 West Java Male 41 2020-09-09 29,897 bp GSAID GH
EPI_ISL_528752 West Java Male 47 2020-09-09 29,881 bp GSAID GH
EPI_ISL_528753 West Java Male 17 2020-09-09 29,868 bp GSAID GH
EPI_ISL_528759 West Java Male 37 2020-09-09 29,865 bp GSAID GH
EPI_ISL_529138 Surabaya Male 63 2020-09-10 29,901 bp GSAID GH
EPI_ISL_529961 Surabaya Male 55 2020-09-10 29,892 bp GSAID L
EPI_ISL_529962 Surabaya Female 43 2020-09-10 29,890 bp GSAID L
EPI_ISL_529963 Surabaya Male 42 2020-09-10 29,891 bp GSAID GH
EPI_ISL_529964 Pasuruan Male 39 2020-09-10 29,894 bp GSAID L
EPI_ISL_529965 Surabaya Female 77 2020-09-10 29,899 bp GSAID GH
EPI_ISL_529966 Sidoarjo Female 40 2020-09-10 29,900 bp GSAID GH
EPI_ISL_529967 Sidoarjo Male 75 2020-09-10 29,902 bp GSAID GH

37
Tabel data tambahan 3. Mutasi SARS-CoV-2 dari sampel Wuhan
Sequence_ID Mutations
EPI_ISL_402120 NS7a P34S
EPI_ISL_402132 Spike F32I
EPI_ISL_406798 NSP3 L1417I, NSP6 N264K
EPI_ISL_406801 NS8 L84S
EPI_ISL_493166 NSP3 W1632stop, NSP10 T51A
hCoV-19_A142 NSP2 N92H, NSP14 P24T
hCoV-19_A148 Spike N856K, NSP2 N92H
hCoV-19_A152 NSP2 N92H
hCoV-19_A160 Spike N856K, NSP2 N92H
hCoV-19_A163 Spike N856K, NSP2 N92H
hCoV-19_A168 NSP2 N92H, NSP12 D62G
hCoV-19_A169 NSP13 V98F
hCoV-19_A170 NSP14 G114D
hCoV-19_A191 M K15Q, NS8 L84S, NSP3 V57L
IPBCAMS-WH-01 NSP3 N1890S, NSP4 F145I
IPBCAMS-WH-03 NSP3 I1426T
IPBCAMS-WH-05 NSP3 G1716V
MN996527 NSP16 D220N
MN996529 NSP3 G1433S, NSP16 K160R
MN996531 NSP3 D1761A, NSP4 T327I

38
Tabel data tambahan 4. Mutasi SARS-CoV-2 dari sampel Indonesia
Sequence_ID Mutations
EPI_ISL_435281 NSP3 S1285F
EPI_ISL_435282 NSP2 I281V, NSP12 A399V
EPI_ISL_435283 Spike T76I, NSP6 L37F
EPI_ISL_437187 Spike D614G, NS3 Q57H, NSP2 T85I, NSP3 Q311R, NSP3 Q941P, NSP12 P323L
EPI_ISL_437188 Spike D614G, Spike Q677H, Spike S116C, NS3 Q57H, NSP12 P227L, NSP12 P323L
EPI_ISL_437189 Spike V622F, NSP12 A399V
EPI_ISL_437191 NSP6 Q208H
EPI_ISL_437192 Spike L822F, Spike T572I, N K347N, NSP12 A399V, NSP12 E744D
EPI_ISL_458079 NSP12 A399V, NSP12 V354L
EPI_ISL_458081 Spike A352S, NSP3 E1271D, NSP13 R392H
EPI_ISL_467374 Spike A672V
EPI_ISL_467376 Spike C1254F
EPI_ISL_511879 Spike D614G, Spike N185Y, NS3 Q57H, NSP3 V950I, NSP6 L37F, NSP12 P227L, NSP12 P323L, NSP15 V172I
EPI_ISL_516800 Spike D614G, NS3 Q57H, NSP3 P822L, NSP12 P323L
EPI_ISL_516806 NSP5 M49I
EPI_ISL_516829 Spike D614G, N Q160R, NS3 A54V, NS3 A99S, NS3 Q57H, NSP3 P679S, NSP12 A656S, NSP12 P323L, NSP13 M576I
EPI_ISL_518819 Spike D614G, Spike S477I, Spike T22P, N P20L, NS3 L106F, NS3 Q57H, NSP3 L27F, NSP12 P227L, NSP12 P323L, NSP15 F15S
EPI_ISL_525492 Spike D614G, NS3 Q57H, NS7a H73Y, NSP3 P822L, NSP12 P323L
EPI_ISL_528745 Spike D614G, NS3 Q57H, NSP2 I273T, NSP12 P227L, NSP12 P323L, NSP14 M500I
EPI_ISL_528746 Spike D614G, NS3 Q57H, NSP2 G339S, NSP6 G277S, NSP12 P227L, NSP12 P323L
EPI_ISL_528747 Spike D614G, N G204R, N R203K, NSP12 P323L, NSP14 A504S
EPI_ISL_528748 Spike D614G, NSP12 P323L
EPI_ISL_528749 Spike D614G, Spike R214L, NS3 Q57H, NSP2 T85I, NSP12 P323L

39
EPI_ISL_528750 Spike D614G, Spike R158S, NS3 Q57H, NS3 T223I, NSP3 A450S, NSP12 A529V, NSP12 P323L, NSP14 A281S
EPI_ISL_528751 Spike D614G, Spike Q677H, NS3 Q57H, NSP4 A446V, NSP12 A529V, NSP12 P227L, NSP12 P323L, NSP16 K160R
EPI_ISL_528752 Spike D614G, N S193I, NS3 Q57H, NSP3 C1641F, NSP3 P822L, NSP3 T64I, NSP12 G596S, NSP12 P323L, NSP15 V127F
EPI_ISL_528753 Spike D614G, NS3 Q57H, NSP2 I273T, NSP12 P227L, NSP12 P323L, NSP14 M500I
EPI_ISL_528759 Spike D614G, Spike Q677H, Spike T95I, NS3 D222Y, NS3 Q57H, NSP12 P227L, NSP12 P323L
EPI_ISL_529138 Spike D614G, NS3 Q57H, NSP12 P323L
EPI_ISL_529961 NSP15 Y225C
EPI_ISL_529963 Spike A67V, Spike D614G, Spike T22I, E P71S, NS3 Q57H, NS8 A51V, NSP3 S1670F, NSP12 P227L, NSP12 P323L
EPI_ISL_529964 Spike A352S, NSP3 E1271D, NSP13 R392H
EPI_ISL_529965 Spike D614G, NS3 Q57H, NSP12 P227L, NSP12 P323L
EPI_ISL_529966 Spike D614G, NS3 Q57H, NSP12 P323L
EPI_ISL_529967 Spike D614G, NS3 Q57H, NSP12 P227L, NSP12 P323L


















