
Capítulo 2 - dragao.co.it.ptdragao.co.it.pt/~fp/tese/Cap2.pdf · descrição clássica da...
72
5 Capítulo 2 Modelos Cocleares 2.1 Introdução Um sistema de pré-processamento espectral baseado nas propriedades conhecidas do sistema auditivo periférico deve começar por simular o funcionamento da cóclea. Assim, neste capítulo é apresentada uma descrição física do funcionamento coclear começando pela descrição clássica da propagação das ondas acústicas ao longo da cóclea. Seguidamente são analisados modelos mais pormenorizados que descrevem a interacção entre as diferentes estruturas no interior da cóclea. São apresentados modelos recentes em que a cóclea é descrita como um sistema activo e com base neles é apresentado um novo modelo coclear referente à cóclea humana. Neste capítulo são também apresentados modelos do ouvido externo e médio, necessários para a completa modelação do ouvido. 2.2 Estrutura e funcionamento da cóclea A cóclea corresponde ao ouvido interno e contém as células responsáveis pela transdução do som. Está embutida numa parte inacessível do crânio totalmente envolvida por osso. Na figura 2.1a) mostra-se o aparelho auditivo humano onde se pode observar a cóclea em
Transcript of Capítulo 2 - dragao.co.it.ptdragao.co.it.pt/~fp/tese/Cap2.pdf · descrição clássica da...

5
Capítulo 2
Modelos Cocleares
2.1 Introdução
Um sistema de pré-processamento espectral baseado nas propriedades conhecidas do
sistema auditivo periférico deve começar por simular o funcionamento da cóclea. Assim,
neste capítulo é apresentada uma descrição física do funcionamento coclear começando pela
descrição clássica da propagação das ondas acústicas ao longo da cóclea. Seguidamente são
analisados modelos mais pormenorizados que descrevem a interacção entre as diferentes
estruturas no interior da cóclea. São apresentados modelos recentes em que a cóclea é
descrita como um sistema activo e com base neles é apresentado um novo modelo coclear
referente à cóclea humana. Neste capítulo são também apresentados modelos do ouvido
externo e médio, necessários para a completa modelação do ouvido.
2.2 Estrutura e funcionamento da cóclea
A cóclea corresponde ao ouvido interno e contém as células responsáveis pela transdução
do som. Está embutida numa parte inacessível do crânio totalmente envolvida por osso. Na
figura 2.1a) mostra-se o aparelho auditivo humano onde se pode observar a cóclea em

6 Capítulo 2 - Modelos Cocleares
forma de espiral com cerca de três voltas, semelhante a uma concha de caracol (daí o nome
de cóclea). O interior da cóclea está seccionado em 3 canais preenchidos por líquido ou
fluido coclear. Estes canais são designados por escalas: a escala vestibular, a escala média e
a escala timpânica. A escala vestibular está separada da escala média por uma membrana
muito fina - a membrana vestibular ou de Reissner - mas que permite a separação de fluidos
cocleares com diferentes composições iónicas. Contudo, em termos hidromecânicos pode
considerar-se a escala vestibular e média como uma única, existindo assim efectivamente
apenas duas escalas: a escala vestibular e a escala timpânica. Estas estão separadas por uma
estrutura designada por partição coclear que compreende a membrana basilar bem como o
órgão de Corti que nela se apoia. É no órgão de Corti que se inserem as células ciliadas,
membrana tectória etc. As duas escalas comunicam entre si apenas numa pequena abertura -
o helicotrema - situada no topo ou ápice da cóclea (ver figura 2.2b).
a) b)
Figura 2.1 - a) - Secção longitudinal do aparelho auditivo humano. EAM: canal auditivo externo;TM - membrana timpânica; OC: cadeia ossicular (ouvido médio); MEC: cavidade do ouvidomédio; OW: janela oval; RW: janela redonda; ET: trompa de Eustáquio; VN, FN, CN: nervosvestibular, facial e coclear. b) - Secção transversal da cóclea. BM: membrana basilar; TM:membrana tectória; RM: membrana vestibular (ou de Reissner); SV: escala vestibular; CC: escalamédia; ST: escala timpânica. De Davis, “Psychophysiology of hearing and deafness”, Wiley,1951, adaptado de [Buser & Imbert, 92].
Na base da cóclea existem ainda duas membranas flexíveis, a janela oval e a janela redonda,
que separam o fluido coclear no interior da cóclea do ar (à pressão atmosférica) existente
no espaço do ouvido médio. A janela oval situa-se na escala vestibular e a janela redonda na
escala timpânica. Estas janelas constituem a ligação do ouvido médio à cóclea estando a
base do estribo apoiada na janela oval. As vibrações do estribo dilatam ou contraem a janela

2.2 Estrutura e funcionamento da cóclea 7
oval e têm um efeito praticamente instantâneo e contrário, tipo “push-pull”, na janela
redonda.
O funcionamento da cóclea pode resumir-se da seguinte forma: as vibrações do tímpano são
transmitidas à cadeia óssea do ouvido médio que funciona como um sistema de alavancas e
que produz uma vibração do estribo tipo vaivém (êmbolo) na janela oval. Os eventos
acústicos que daqui resultam podem ser vistos como duas ondas. A vibração na janela oval
produz variações de pressão no fluido coclear que se propagam pela escala vestibular,
passam pelo helicotrema e produzem uma vibração simétrica na membrana da janela
redonda. Esta onda propaga-se à velocidade do som em líquidos e não produz forças na
partição coclear pois é a mesma em ambas as escalas. Além disso, a velocidade de
propagação desta onda pode considerar-se infinita o que significa admitir que o fluido
coclear é incompressível. Contudo, a existência desta onda de pressão vai gerar um
gradiente de pressões através da partição coclear. É esta diferença de pressões que vai
exercer uma força na partição pondo-a em movimento, movimento este que também se
propaga, agora mais lentamente, ao longo da cóclea. O deslocamento da partição por sua
vez vai fazer com que os cílios das células ciliadas sejam deflectidos o que produz uma
variação de tensão através das membranas das células ciliadas. Esta transdução mecano-
eléctrica vai produzir os disparos nas fibras do nervo coclear, transmitindo assim a
informação acústica a centros neuronais mais elevados da via auditiva.
janela oval
janela redonda
estribo
escala vestibular escala timpânica
estribo
janela redonda
partição coclear
escala vestibular
escala timpânica helicotrema
P(x)
a) b)
Figura 2.2 - a) Esquema da cóclea. b) Esquema simplificado com a cóclea “desenrolada”. Avibração do estribo cria variações de pressão no fluido coclear; as diferenças de pressão através dapartição produzem uma onda que se propaga ao longo da cóclea.
As características físicas da partição coclear vão ditar as propriedades da onda de gradiente
de pressão que se propaga ao longo da cóclea. Uma vez que a rigidez da partição decresce

8 Capítulo 2 - Modelos Cocleares
desde a base (janelas) até ao ápice (helicotrema), a velocidade de propagação também
decresce tornando-se o comprimento de onda cada vez menor para uma dada frequência de
excitação: λ=v/f (fig. 2.2b). A onda é inicialmente “longa” tornando-se progressivamente
mais “curta”. Como resultado deste encurtamento a onda torna-se dispersiva e extingue-se
numa região curta. A amplitude máxima atinge-se numa posição chamada posição
característica que é função da frequência de excitação. Por outro lado, cada localização da
cóclea tem associada uma frequência característica (CF). Estas frequências estão mapeadas
na cóclea de acordo com um mapa posição-frequência a que se dá o nome de tonotopia. As
frequências características decrescem desde a base da cóclea até ao ápice da mesma forma
que a rigidez da partição decresce ao longo da sua extensão. Ondas com alta frequência
extinguem-se perto da base enquanto ondas de mais baixa frequência penetram mais
profundamente na cóclea. Este funcionamento hidromecânico em termos de ondas
progressivas (“traveling waves”) e a tradução posição-frequência constitui a base da análise
espectral operada pela cóclea.
Por último convém referir que as vibrações cocleares são de dimensões moleculares, da
ordem dos nanometros, e que a cóclea pode detectar uma variação de pressão de 20µPa
que corresponde a 2x10-10 da pressão atmosférica normal [Buser & Imbert, 92]. Apesar
desta extrema sensibilidade, a cóclea pode suportar, por pequenos períodos, pressões
sonoras 1 milhão de vezes superiores em amplitude que a pressão do limiar de audição. Tal
gama dinâmica (120dB) é possível devido a uma redução de sensibilidade mecânica à
medida que o nível sonoro cresce. Daqui resultam os termos controlo automático de ganho
(AGC) e não-linearidade compressiva, relativos às respostas cocleares em função do nível
de excitação. Para baixos níveis de excitação a cóclea comporta-se de uma forma
aproximadamente linear e portanto os modelos cocleares são válidos apenas para esta
situação.
A vibração da partição (membrana basilar e órgão de Corti) relativa às estruturas ósseas e
fluido circundante é conhecida como vibração macromecânica enquanto a complexa
vibração das partes constituintes do órgão de Corti relativas umas às outras é conhecida
como vibração micromecânica. Os modelos cocleares seguem a mesma classificação,
falando-se de modelos macromecânicos e modelos micromecânicos da cóclea.

2.2 Estrutura e funcionamento da cóclea 9
2.2.1 Modelação coclear
Seguindo um percurso clássico, a análise coclear pode ser feita em termos do
comportamento mecânico da membrana basilar e do fluido envolvente, tendo em conta as
suas propriedades físicas e geométricas, (macromecânica), e depois em termos do
funcionamento do órgão de Corti e das células ciliadas (micromecânica). A modelação
coclear começou por considerar as propriedades mecânicas da cóclea onde a partição
coclear, constituída pela membrana basilar e pelo órgão de Corti nela inserida, vibrava em
uníssono e sem interacção entre estas duas estruturas. Os modelos a uma dimensão (1D)
apresentavam resultados encorajadores, contudo pensou-se que a discrepância em relação
aos dados experimentais se devia à simplicidade do modelo e começou a investigar-se
modelos a duas e três dimensões, embora sem drásticas melhorias em relação ao modelo
1D. Foi principalmente a descoberta das emissões otoacústicas que veio alterar e alargar a
visão sobre o funcionamento da cóclea. De facto, um sistema que espontaneamente produz
vibrações acústicas (de dentro para fora, neste caso) não poderia ser um sistema puramente
passivo. Presentemente a modelação coclear passou a incidir principalmente na
micromecânica, ou seja, a considerar a interacção da membrana tectória e das células
ciliadas com a membrana basilar, e menos na dimensionalidade do modelo. Apesar de
existirem ainda muitas incógnitas, os modelos cocleares recentes conseguem descrever com
algum rigor os dados experimentais.
Existe hoje crescente evidência de que as células ciliadas externas (CCEs) são responsáveis
pela elevada selectividade das respostas cocleares a baixos níveis de excitação. Esta
evidência resulta principalmente de dois factos: o primeiro é que, em cócleas vivas onde as
CCEs são destruídas (por exemplo por medicamentos ototóxicos) ou em cadáveres, as
respostas cocleares apresentam uma selectividade muito inferior à situação de
funcionamento normal. O outro facto é a recente descoberta da capacidade de alongamento
das CCEs por excitação bioeléctrica. Esta descoberta foi feita com experiências in vitro por
cultura de CCEs, mas significa que estas células podem produzir forças que vão alterar os
padrões acústicos ao longo da cóclea.
Inicialmente a suposição de que a cóclea seria um sistema activo conduziu à teoria do
“amplificador coclear” [Kolston et al, 90]). Mais tarde mostrou-se, tendo como base os
dados mecânicos obtidos experimentalmente, que um sistema puramente passivo não
poderia produzir as curvas observadas [de Boer 83, 91].

10 Capítulo 2 - Modelos Cocleares
Apesar de alguma controvérsia à volta da hipótese de “actividade” coclear (que ainda não
está de todo resolvida), a verdade é que praticamente todos os modelos cocleares recentes
passaram a considerar a cóclea como um sistema activo, pelo menos numa região perto do
ponto de máxima selectividade.
2.3 Modelo macromecânico a uma dimensão
Um modelo matemático da cóclea pode ser desenvolvido considerando as suas propriedades
físicas e fisiológicas. Para que o modelo seja matematicamente tratável, algumas
simplificações deverão ser feitas. Uma destas simplificações diz respeito à geometria da
cóclea. Uma vez que o raio de curvatura da espiral coclear é muito superior ao
comprimento de onda das ondas cocleares, pode considerar-se a cóclea “desenrolada” sem
que isso afecte significativamente as respostas [Patuzzi, 96]. Por outro lado, os dois canais
cocleares podem considerar-se geometricamente simples com secção rectangular ou
circular. Por último, a área destas secções pode considerar-se constante dando origem a um
modelo cilíndrico ou paralelipípedo (“box model”, fig. 2.3).
Outra simplificação diz respeito ao acoplamento longitudinal da partição coclear: considera-
se que pontos da partição em diferentes posições longitudinais não têm acoplamento
mecânico directo a não ser indirectamente através do fluído coclear [de Boer, 91], ou seja,
cada segmento (infinitesimal) da partição vibra independentemente de qualquer outro. Isto
significa que partição coclear pode ser descrita pela sua impedância mecânica local (ou
pontual). Esta simplificação veio a mostrar-se ser essencialmente correcta1 e é feita na
grande maioria dos modelos cocleares propostos até hoje.
Uma outra simplificação consiste em admitir que o comprimento de onda do gradiente de
pressão coclear é muito superior às dimensões da secção transversal do modelo. Desta
forma pode considerar-se que os efeitos cinéticos dos movimentos do fluído coclear só se
fazem sentir numa única direcção: a direcção longitudinal, x (fig. 2.3), denominada posição
ou distância coclear. Esta análise conduz à definição de um modelo a uma dimensão
(modelo 1D) também geralmente referido como modelo de “ondas longas” (“long wave
1 O acoplamento longitudinal entre regiões adjacentes da partição coclear veio a mostrar-se ser fraco.[Patuzzi, 96]. Apesar do nome, a membrana basilar funciona de uma forma mais parecida com um xilofonedo que com uma membrana elástica ou corda de piano.
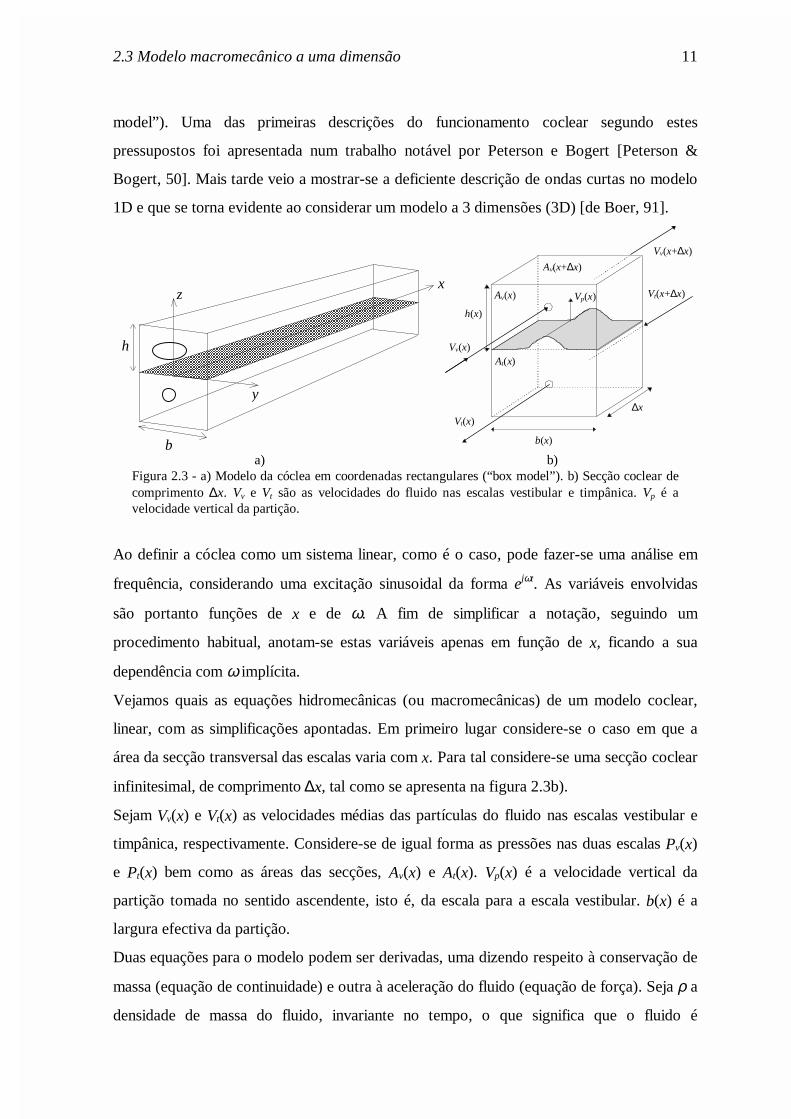
2.3 Modelo macromecânico a uma dimensão 11
model”). Uma das primeiras descrições do funcionamento coclear segundo estes
pressupostos foi apresentada num trabalho notável por Peterson e Bogert [Peterson &
Bogert, 50]. Mais tarde veio a mostrar-se a deficiente descrição de ondas curtas no modelo
1D e que se torna evidente ao considerar um modelo a 3 dimensões (3D) [de Boer, 91].
h
y
zx
b
h(x)
b(x)
∆x
Vp(x)
Vv(x)
Vv(x+∆x)
Vt(x)
Vt(x+∆x)Av(x)
At(x)
Av(x+∆x)
a) b)Figura 2.3 - a) Modelo da cóclea em coordenadas rectangulares (“box model”). b) Secção coclear decomprimento ∆x. Vv e Vt são as velocidades do fluido nas escalas vestibular e timpânica. Vp é avelocidade vertical da partição.
Ao definir a cóclea como um sistema linear, como é o caso, pode fazer-se uma análise em
frequência, considerando uma excitação sinusoidal da forma ejωt. As variáveis envolvidas
são portanto funções de x e de ω. A fim de simplificar a notação, seguindo um
procedimento habitual, anotam-se estas variáveis apenas em função de x, ficando a sua
dependência com ω implícita.
Vejamos quais as equações hidromecânicas (ou macromecânicas) de um modelo coclear,
linear, com as simplificações apontadas. Em primeiro lugar considere-se o caso em que a
área da secção transversal das escalas varia com x. Para tal considere-se uma secção coclear
infinitesimal, de comprimento ∆x, tal como se apresenta na figura 2.3b).
Sejam Vv(x) e Vt(x) as velocidades médias das partículas do fluido nas escalas vestibular e
timpânica, respectivamente. Considere-se de igual forma as pressões nas duas escalas Pv(x)
e Pt(x) bem como as áreas das secções, Av(x) e At(x). Vp(x) é a velocidade vertical da
partição tomada no sentido ascendente, isto é, da escala para a escala vestibular. b(x) é a
largura efectiva da partição.
Duas equações para o modelo podem ser derivadas, uma dizendo respeito à conservação de
massa (equação de continuidade) e outra à aceleração do fluido (equação de força). Seja ρ a
densidade de massa do fluido, invariante no tempo, o que significa que o fluido é

12 Capítulo 2 - Modelos Cocleares
incompressível. A massa de fluido por unidade de tempo que entra numa secção coclear
infinitesimal através da área Av(x) da escala vestibular é ρVv(x)Av(x). Esta massa somada à
massa de fluido deslocada (para cima) pela vibração da partição através da área da partição
Ap(x)=b(x)∆x, deverá ser igual à massa que sai através da área Av(x+∆x):
ρ ρ ρ V x x A x x V x A x V x b x xv v v v p( ) ( ) ( ) ( ) ( ) ( )+ + = +∆ ∆ ∆ .
Uma expressão idêntica aplica-se à escala timpânica. Quando ∆x tende para zero resultam
as seguintes equações diferenciais:
( )d
dxV x A x b x V xv v p( ) ( ) ( ) ( )= , (2.1a)
( )d
dxV x A x b x V xt t p( ) ( ) ( ) ( )= . (2.1b)
Por outro lado, a variação de pressão nas escalas vai acelerar o fluido. Para uma coluna de
fluido invíscido de largura ∆x, teremos, aplicando a 2ª lei de Newton [de Boer, 80],
[Koshigoe et al, 83]:
( )d
dxP x A x x A x x j V xv v v v( ) ( ) ( ) ( ( ))∆ ∆≅ − ρ ω .
A massa de fluido na secção coclear não é exactamente ρAv∆x daí a aproximação na
expressão anterior. Uma expressão mais exacta pode ser obtida utilizando a equação de
força ou equação de Euler linearizada [Kinsler et al, 82]: ∇ p = −ρdv/dt,2 que se obtém
considerando um elemento de volume infinitesimal. Se as variáveis são definidas apenas a
uma dimensão, a equação de Euler aplicada à escala vestibular é a seguinte:
( )dP x
dxj V x
j
A xV x A xv
vv
v v
( )( )
( )( ) ( )= − = −ωρ
ωρ .
O produto de uma velocidade linear por uma área é usualmente referida como velocidade
volúmica (devido à sua dimensão: m3/s) e será designada por U(x). Considerando a
viscosidade, as equações para as duas escalas são as seguintes:
dP x
dx
j Q x
A xU xv v
vv
( ) ( )
( )( )= −
ωρ , (2.2a)
dP x
dx
j Q x
A xU xt t
tt
( ) ( )
( )( )= +
ωρ . (2.2b)
Qv(x) e Qt(x) são funções que tomam em conta as contribuições resistivas e reactivas da
densidade de massa associadas ao arrastamento viscoso do fluido. Para o caso de um fluido

2.3 Modelo macromecânico a uma dimensão 13
invíscido, teremos Q(x)=1, caso contrário, Q(x) apresenta um termo que tende a ser
inversamente proporcional à raiz quadrada da frequência3.
Em adição às relações anteriores, é necessário ainda uma expressão para o movimento da
partição. Esta relação obtém-se através da impedância da partição. A impedância específica
da partição coclear é, por definição, a relação entre a diferença de pressões através da
partição, da escala timpânica para a escala vestibular, com a sua velocidade pontual vertical:
Z xP x P x
V xpt v
p
( )( ) ( )
( )=
− . (2.3)
Assume-se com esta equação que um elemento b(x)∆x da superfície da partição não tem
acoplamento mecânico com os elementos vizinhos.
A análise do modelo fica extremamente simplificada se se igualar as áreas das secções
vestibular e timpânica, o que se mostra ser aproximadamente correcto excepto na base da
cóclea. De facto, definindo a diferença de pressões através da partição,
P x P x P xv t( ) ( ) ( )= − , (2.4)
e a média das velocidades volúmicas nas duas escalas por,
( )U x U x U xv t( ) ( ) ( )= +1
2, (2.5)
então, com Av(x)=At(x)= A(x), resulta Qv(x)=Qt(x)=Q(x), e o número de equações reduz-se
a metade, vindo:
d
dxU x b x V x
P x A x
h x Z xpp
( ) ( ) ( )( ) ( )
( ) ( )= = − , (2.6a)
dP x
dx
j Q x
A xU x
( ) ( )
( )( )= −
2 ωρ. (2.6b)
Na equação (2.6a) h(x) é a altura efectiva da escala, a relação entre a área A(x) e a largura
da partição b(x). Conjugando as últimas duas expressões resulta a equação diferencial de 2ª
ordem do modelo:
( )d P x
dx
d
dxQ x A x
dP x
dx
j Q x
h x Z xP x
p
2
2
2( )log ( ) ( )
( ) ( )
( ) ( )( )= +
ωρ . (2.7)
2 As letras minúsculas, p e v, indicam pressão e velocidade em função do tempo.3 Para o caso de um modelo em que as áreas das secções são circulares, ( )Q x J v vJ v( ) ( ) ( ( ))= −1 1 2 1 0
,
v jf A x= −2 ρ µ( ) , onde Jn(v) é a função de Bessel de 1ª espécie, ordem n e onde µ é o coeficiente de
viscosidade [Koshigoe et al. 83], [Puria & Allen, 91]. Para v>10, Q x j fA x( ) ( ) ( )≈ + −1 1 µ ρ .

14 Capítulo 2 - Modelos Cocleares
Vejamos agora duas simplificações usuais na modelação coclear. Uma vez que o efeito da
viscosidade do fluido só se faz sentir a baixas frequências, esta propriedade é muitas vezes
ignorada, isto é, toma-se Q(x)=1. Além disso, se a área das secções for constante com x,
como é o caso de um modelo paralelipípedo ou cilíndrico, a equação anterior simplifica-se
resultando na seguinte expressão:
d P x
dx
j
hZ xP x
p
2
2
2( )
( )( )= ⋅
ωρ . (2.8)
Nas secções seguintes iremos analisar o modelo paralelipípedo descrito por esta equação.
Antes de mais, é interessante verificar que esta equação, com Zp constante, é
frequentemente encontrada em vários fenómenos físicos ondulatórios, nomeadamente, na
propagação de ondas planas e na análise de linhas de transmissão de sinais eléctricos (linhas
homogéneas). No caso presente, a impedância da partição varia muito significativamente
com a distância à base da cóclea, x. A cóclea funciona assim analogamente a uma linha de
transmissão não-homogénea. A constante de propagação, o número de onda, a velocidade
de fase, etc., passam neste caso a ser função de x.
No caso de uma impedância da partição constante (apesar de incorrecto no caso da cóclea),
a solução da equação anterior seria:
P x c e c ex x( ) = +− +1 2
γ γ (2.9)
onde c1 e c2 são constantes e γ é a constante de propagação, dada por:
γωρ
α β= = = +2 j
hZjk j
p
. (2.10)
A interpretação desta solução é a existência de uma onda (plana) progressiva (sinal −), com
coeficiente de atenuação α e coeficiente de fase β; e uma onda em direcção contrária (sinal
+), que é o resultado acumulado de reflexões. A variável k é denominada número de onda.
O comprimento de onda é λ=2π/β, embora seja usual considerar o comprimento de onda
complexo, através do número de onda: λc=2π/k . As constantes c1 e c2 dependem das
impedâncias de “fonte” e de “terminação” (impedância do ouvido médio e impedância do
helicotrema, no caso da cóclea).
A analogia com linhas de transmissão consegue-se associando P(x) à tensão na linha de
transmissão e U(x) à corrente. Podem assim definir-se a impedância distribuída (em série) e
a admitância distribuída (em paralelo). De acordo com as variáveis em questão, estas

2.3 Modelo macromecânico a uma dimensão 15
impedâncias são impedâncias acústicas, a relação entre pressão e velocidade volúmica4. A
impedância e admitância distribuídas são, considerando as equações (2.6), as seguintes:
Z xj Q x
A xser ( )( )
( )=
2 ωρ, (2.11a)
Y xh x Z x A xpar
p
( )( ) ( ) ( )
=1
. (2.11b)
No caso de um modelo com A(x) e Q(x) constantes, o modelo pode ser definido em termos
de pressão e velocidade pontual, reduzindo-se as equações (2.6) às seguintes:
d
dxV x
P x
hZ xp
( )( )
( )= − , (2.12a)
d
dxP x j V x( ) ( )= −2 ωρ . (2.12b)
Os parâmetros distribuídos são, neste caso, impedâncias específicas por unidade de
comprimento:
Z jser = 2 ωρ , (2.13a)
Y xh Z xpar
p
( )( )
=⋅
1 . (2.13b)
Através dos parâmetros distribuídos definem-se ainda duas outras grandezas, a impedância
característica da linha e o número de onda. No caso de uma linha homogénea estes
parâmetros são fixos e caracterizam completamente a linha. No presente caso, estes
parâmetros são funções de x. A solução da equação de onda não fica determinada através
deles e são agora referidos como “locais” [de Boer, 83]:
Z x Z x Y xloc ser par0 ( ) ( ) ( )= , (2.14a)
jk x Z x Y xloc ser par( ) ( ) ( )= , (2.14b)
uma vez que dão uma indicação da solução se a linha fosse homogénea e definida com os
valores dessa localização particular x.
A impedância da partição varia substancialmente com x, como foi referido, principalmente
devido à rigidez da partição que é dura na base e muito mais flexível junto ao ápice
4 De acordo com as variáveis mais adequadas na análise acústica, pressão ou força e velocidade linear ouvelocidade volúmica, assim são definidos três tipos de impedância:
Impedância específica: Z(s)=P/V [N.m-3.s] ou [Kg.m-2.s-1]Impedância acústica: Z(a)=P/U [N.m-5.s] ou [Kg.m-4.s-1]Impedância mecânica: Z(m)=F/V [N.m-1.s] ou [Kg.s-1]

16 Capítulo 2 - Modelos Cocleares
(helicotrema) sendo esta variação de algumas ordens de grandeza. É aliás esta
particularidade que confere à cóclea uma grande selectividade em frequência, envolvendo
uma gama de frequências audíveis de mais de 9 oitavas [Boer, 83, 91]. As impedâncias
distribuídas são portanto função da posição x. A solução da equação do modelo é do
mesmo tipo que a equação (2.9) mas com uma constante de propagação (ou número de
onda) a variar com a posição x.
Não existe uma solução analítica simples e genérica para a equação do modelo, a não ser
com simplificações drásticas. Na secção seguinte irão ser abordados métodos numéricos de
resolução do modelo.
2.3.1 Solução numérica
A única forma de resolver a equação (2.7) ou (2.8) sem restrições é através de métodos
numéricos. No desenvolvimento seguinte iremos considerar o caso mais simples da equação
(2.8). Um método óbvio e simples consiste em concatenar secções infinitesimais com os
parâmetros distribuídos definidos. Este método equivale a considerar aproximações de
primeira ordem para as derivadas de P(x) e V(x):
dP x
dx
P x x P x
x
( ) ( ) ( )≈
+ −∆∆
, (2.15)
dV x
dx
V x V x x
x
( ) ( ) ( )≈
− − ∆∆
, (2.16)
onde ∆x corresponde ao comprimento infinitesimal de uma secção coclear. Através das
equações anteriores e das equações (2.12) e (2.13), obtemos as seguintes relações para uma
secção infinitesimal:
P x P x x j xV x Z x V xser( ) ( ) ( ) ( )− + = = ⋅∆ ∆ ∆2 ωρ , (2.17)
V x x V xP x
hZ xx Y x x P x V x
ppar par( ) ( )
( )
( )( ) ( ) ( )− − = = ⋅ =∆ ∆ ∆ . (2.18)
O circuito eléctrico equivalente que verifica estas equações encontra-se na figura 2.4. Trata-
se portanto do circuito eléctrico equivalente de uma secção coclear infinitesimal com
parâmetros distribuídos Zser e Ypar(x) (ou com parâmetros concentrados zser=Zser∆x,
ypar(x)=1/zpar(x)=Ypar(x)∆x). A velocidade Vpar(x) relaciona-s com a velocidade da partição
através da relação Vp(x)=hVpar(x).

2.3 Modelo macromecânico a uma dimensão 17
V(x-∆x)
Zser∆x Zser∆x
P(x-∆x) P(x+∆x)Zpar(x)∆x
V(x)
P(x)
Vpar(x)
Figura 2.4 - Esquema eléctrico de uma secção coclear infinitesimal.
Para resolver a equação (2.8) é necessário considerar condições fronteira. No presente caso,
conhecida a impedância do ouvido externo e médio, Zm, poderá considerar-se a pressão
externa ou a pressão timpânica como excitação de entrada, ficando assim definida uma das
condições fronteira5. A outra condição fronteira implica a definição da impedância do
helicotrema, Zh, onde o fluido coclear comunica entre as duas escalas. Nesta situação, pode
considerar-se, sem grande erro, que a diferença de pressões entre as duas escalas é nula, ou
seja, P(x=L)=0, onde L corresponde ao comprimento coclear. Isto equivale a considerar a
impedância do helicotrema nula.
Discretizando a cóclea em N secções de comprimento ∆x e designando P(n) como sendo a
diferença de pressão P(x=n∆x), n=0..N, onde ∆x=L/N, a equação diferencial do modelo
resulta na seguinte equação de diferenças:
P nz
z nP n P nser
par
( )( )
( ) ( )− − +
+ + =1 2 1 0 . (2.19)
O modelo assim definido corresponde ao circuito eléctrico da figura 2.5. Através desta
figura facilmente se obtém a equação de fronteira (para n=1):
P P
Z
P P
zi
m ser
−=
−( ) ( ) ( )0 0 1. (2.20)
5 É também usual definir as equações do modelo em função da velocidade do estribo normalizada (ouV(0)=1). Desta forma evita-se a definição da impedância do ouvido externo e médio para avaliar o modelococlear. A “fonte” a considerar no modelo deverá ser o equivalente de Thévenin do conjunto ouvido médio eexterno reduzido à entrada da cóclea.

18 Capítulo 2 - Modelos Cocleares
zpar(1) zpar(k)Pi
Zm P(0) P(1) P(k) P(N-1)
zpar(N-1)
zser(1)
Zr(k)
P(N)
V(0) V(k-1) V(N-1)
...
... ...
...
Zh
zser(2) zser(k) zser(N)
V(k)
Zr(k-1)
Figura 2.5 - Modelo equivalente de linha de transmissão. Zm e Pi correspondem ao equivalente deThévenin do ouvido externo e médio referido à entrada da cóclea. Zh é a impedância do helicotrema.Zr(k) é a impedância vista à direita da secção coclear k, Zr(k)=P(k)/V(k). As impedâncias zser=Zser∆xe zpar=Zpar/∆x são definidas através das equações (2.11) ou (2.13). No caso das equações (2.11) deveconsiderar-se a velocidade volúmica U(k) em vez da velocidade pontual V(k).
Para n=N−1, considerando a impedância do helicotrema nula, teremos P(N)=0. Assim, as
equações (2.19) e (2.20) definem um sistema de N equações a N incógnitas, P(n),
n=0,..,N−1, sendo a matriz do sistema tridiagonal. A solução deste sistema de equações
pode ser obtida por eliminação gaussiana ([de Boer, 80], [Neely & Kim, 86]). Existem
contudo outras formas alternativas de resolver este sistema de equações utilizando a teoria
da análise de circuitos eléctricos.
Uma destas formas consiste em utilizar parâmetros de transmissão, método utilizado
principalmente pelo grupo de Allen ([Puria & Allen, 91], [Allen & Fahey, 93], [Allen &
Neely, 92]).
Na análise com parâmetros de transmissão define-se, para uma secção coclear, o seguinte
sistema:
P n
V n
A B
C D
P n
V n
P n
V nn n
n nn
( )
( )
( )
( )
( )
( )
−−
=
⋅
= ⋅
1
1T . (2.21)
No caso presente, os elementos da matriz de transmissão são: An=1+zser(n)ypar(n);
Bn=zser(n); Cn=ypar(n) e Dn=1. As matrizes Tn têm determinante unitário (propriedade da
reciprocidade do circuito), pelo que se pode considerar o sistema inverso. Cascatas de
secções cocleares são representadas por multiplicações das matrizes de transmissão. Assim,
por exemplo, aplicando várias vezes a definição anterior, resulta, uma vez que V(N)=0,
P n
V n
P N P
Vkk n
N
kk
n( )
( )
( ) ( )
( )
=
⋅
=
⋅
= + =
−
∏ ∏T T1 1
1
0
0
0 . (2.22)
Definindo a matriz produto da seguinte forma (com i < j):

2.3 Modelo macromecânico a uma dimensão 19
T Ti j nn i
ji j i j
i j i j
A B
C D,
, ,
, ,
= =
=∏ , Ti j
i j i j
i j i j
D B
C A,
, ,
, ,
− =−
−
1 , (2.23)
resulta que, qualquer pressão, velocidade ou impedância pode ser determinada, admitindo
uma impedância do helicotrema não nula. Seguem-se alguns exemplos:
P A P N
V C P N
P n A P NA
AP D P C V
ZP
V
A
C
N
N
n N
n N
Nn n
i
N
N
( ) ( )
( ) ( )
( ) ( ) ( ) ( ) ( )
( )
( )
,
,
,
,
,, ,
,
,
0
0
0 0 0
0
0
1
1
1
1
11 1
1
1
==
= = = −
= =
++
.
(2.24)
Trata-se de um método elegante, contudo com problemas de gama numérica uma vez que
os elementos da matriz Ti,j, podem ter expoentes, em módulo, muito elevados. Por
exemplo, P(N) apresenta, para frequências de excitação não muito baixas, um valor
praticamente nulo pelo que A1,N terá de apresentar um valor extremamente elevado.
Uma outra forma de resolver o sistema de equações consiste em calcular em primeiro lugar
as impedâncias equivalentes, Zr(k), vistas à direita de cada secção da linha6 (ver figura 2.5).
Fazendo inicialmente este cálculo desde o helicotrema até à base, podem seguidamente
calcular-se as pressões P(n). De acordo com a figura 2.5, Zr(N-1)= zser(N)+Zh e a definição
recorrente de Zr é :
Z k z k z k Z k k Nr ser par r//( ) ( ) ( ) ( ) ,..,− = + = −1 2 0 , . (2.25)
Uma vez definida esta impedância, podemos calcular a função de transferência para cada
secção k, utilizando a definição anterior:
P k
P k
z k Z k
z k Z k z k
Z k x
Z kpar r
par r ser
ser
r
////
( )
( )
( ) ( )
( ) ( ) ( )
( )
( )−=
+= −
−11
1
∆. (2.26)
A diferença de pressão para a secção k em função da diferença de pressão nas janelas é
simplesmente o produto destas funções de transferência:
P k
P
Z n x
Z nser
rn
k( )
( )
( )
( )01
11
= −−
=∏ ∆
. (2.27)
6 Considera-se neste caso zser variável com x, o que torna as equações (2.25) a (2.27) válidas para ummodelo definido com impedâncias acústicas e com áreas da secções cocleares não constantes.

20 Capítulo 2 - Modelos Cocleares
Esta solução é numericamente estável, embora poucas vezes referenciada na literatura. Uma
das referências onde esta solução é explicitamente indicada é em [Kolston et al, 90].
Esta abordagem tem ainda a vantagem de evidenciar a solução exacta para o modelo em
função de Zr. De facto, com ∆x muito pequeno, obtemos a seguinte aproximação:
( ) ( )ln ( ) ln ( ) ln ( ) ( ) ln ( ) ( ) ( )P k P Z n x Z n P Z n Z n xser rn
k
ser rn
k
= + − − ≈ − −= =
∑ ∑0 1 1 0 11 1
∆ ∆
e ao fazer ∆x tender para zero, o somatório anterior tende para um integral, resultando a
seguinte expressão para P(x) 7:
P x PZ x
Z xdxser
r
x
( ) ( ) exp( )
( )= −
∫00
. (2.28)
Esta forma exacta resulta da equação (2.12b) e da definição de Zr(x):
Z xP x
V xr ( )( )
( )= ; (2.29a)
dP x
dxP x Z x V xser
( )( ) ( ) ( )= ′ = − . (2.29b)
A equação resultante é:
′
= −P x
P x
Z x
Z xser
r
( )
( )
( )
( ) , (2.30)
que tem como solução precisamente a equação anteriormente referida. O problema reside
na determinação da impedância Zr(x). Introduzindo a definição de Zr(x) na equação (2.8),
chegamos, depois de algumas manipulações algébricas, à seguinte equação diferencial:
dZ x
dx
Z x
Z xZ xr r
parser
( ) ( )
( )( )= −
2
, (2.31)
que é (para Zser constante) uma equação de Ricatti [Kaernbach et al, 86] e para a qual não
existe também uma solução analítica genérica. Existem trabalhos onde uma análise criteriosa
desta impedância é feita com o fim de analisar as reflexões em modelos cocleares
[Viergever & de Boer, 87], [de Boer, 91].
Embora não exista uma solução analítica genérica para o modelo, existe, contudo, uma
solução aproximada, utilizada desde há muito em mecânica quântica e conhecida por
solução LG (Liouville-Green) ou solução WKB, ou ainda, solução WKBJ (que deriva dos
7 O número de secções cocleares a considerar deve ser suficientemente grande. Uma forma empírica dedeterminar o número adequado de secções consiste em considerar sucessivamente um maior número desecções até que o resultado obtido não difira significativamente do resultado obtido na iteração anterior.

2.3 Modelo macromecânico a uma dimensão 21
autores Wentzel, Kramers, Brillouin e Jeffreys) [Allen, 77], [de Boer, 80]. Esta solução
parte das impedâncias conhecidas, neste caso Zser e Zpar, admitindo uma solução genérica
para uma onda progressiva da seguinte forma, onde k(x) é o número de onda:
P x P jk x dxx
( ) ( ) exp ( )= ⋅ −
∫0
0
. (2.32)
Comparando esta expressão com a equação (2.28), chega-se à conclusão que o número de
onda está relacionado com Zr da seguinte forma:
jk xZ x
Z xser
r
( )( )
( )= . (2.33)
Por outro lado, comparando esta expressão com o número de onda local (ver (2.14b)),
jk xZ x
Z xlocser
par
( )( )
( )= . (2.34)
verifica-se que estas duas expressões só serão idênticas se Zr=(ZserZpar)1/2=Z0loc(x). Isto é,
para o caso de uma linha homogénea ou para o caso da impedância da partição variar muito
lentamente com x, a impedância característica local e o número de onda local são uma boa
aproximação dos valores verdadeiros.
A solução LG é mais elaborada partindo da seguinte aproximação quanto à derivada do
número de onda: k'(x)=k'loc(x). Utilizando esta aproximação bem como a equação (2.31),
depois de algumas operações algébricas, chega-se à seguinte relação:
k x k x jk x
k xk x
j k x
k xlocloc
locloc
loc
loc
( ) ( )( )
( )( )
( )
( )≅ −
′≈ −
′1
22 . (2.35)
A última aproximação é apenas válida na seguinte condição:
′ <<k kloc loc2 , (2.36)
condição esta que é precisamente a condição de validade da solução LG. Finalmente,
integrando jk(x) de acordo com (2.32), chegamos à solução aproximada LG:
P x Pk
k xj k x dxloc
locloc
x
( ) ( )( )
( )exp ( )≅ ⋅ −
∫0
01 1
0
. (2.37)
Esta solução aproximada é extensamente referida na literatura sobre modelos a uma
dimensão. A vantagem da existência de uma expressão analítica, mesmo que aproximada, é
permitir clarificar a dependência das respostas cocleares em função dos vários parâmetros
físicos. Esta expressão é especialmente útil para baixos valores de x onde o 2º factor da
expressão anterior varia mais lentamente que o 1º. Tal será verificado mais adiante quando

22 Capítulo 2 - Modelos Cocleares
se falar na determinação da impedância de entrada da cóclea. Contudo, para os casos de
interesse abordados neste trabalho, especialmente no caso de impedâncias activas, a
condição (2.36) não se verifica (pelo menos numa determinada gama de valores de x
quando a impedância da partição varia acentuadamente).
Para finalizar convém notar que, em qualquer dos métodos numéricos de resolução
apontados, a solução obtida contém a contribuição da onda incidente bem como toda e
qualquer reflexão que exista na linha por desadaptação.
2.3.2 Impedância da partição - Modelo clássico
Até ao momento foram indicadas as equações macromecânicas de um modelo coclear onde
a impedância da partição joga um papel fundamental, embora esta impedância não tivesse
sido definida. A impedância da partição depende das propriedades da membrana basilar bem
como das propriedades do órgão de Corti. Seguindo a sequência temporal da modelação
coclear, vamos em primeiro lugar considerar o caso em que a partição coclear corresponde
simplesmente à membrana basilar onde o órgão de Corti nela apoiada não interfere
movendo-se solidariamente. O modelo assim definido será referido como modelo clássico.
Posteriormente serão analisados modelos micromecânicos que implicam uma definição mais
elaborada da impedância da partição.
Para definir uma impedância da partição com o pressuposto atrás definido, basta considerar
3 parâmetros mecânicos específicos (por unidade de área): massa M, resistência, R e
rigidez, S. Com s=jω sendo ω a frequência angular de excitação, a impedância específica da
partição toma a seguinte forma:
Z x sM x R x S x sp ( ) ( ) ( ) ( )= + + . (2.38)
Apesar de existirem algumas variações na definição da dependência dos parâmetros com x,
a abordagem mais usual consiste em considerar a massa da partição constante e a rigidez a
variar de forma exponencial. Além disso, a resistência R(x) mostra-se importante apenas na
região de ressonância (onde as outras duas componentes se anulam), e, por conveniência,
muitas vezes considera-se também a variar de forma exponencial. Sendo então a rigidez
definida na forma S(x)=S0e-ax, com S0 a rigidez da partição na base da cóclea, a frequência
de ressonância da membrana é:

2.3 Modelo macromecânico a uma dimensão 23
ω ω0 02
02( ) ( ) / /x S x M S M e ea x
ca x= = =− − . (2.39)
Esta relação traduz a tonotopia coclear, isto é, relaciona a frequência de ressonância com a
posição coclear, relação esta que é exponencial. Sabe-se que esta relação exponencial é
essencialmente correcta para posições basais [Békési, 60], embora menos correcta para
posições apicais. Os estudos sobre o nervo coclear [Liberman, 82], apontam
consistentemente para uma sintonização aproximadamente exponencial das fibras do nervo
coclear de acordo com a sua distribuição ao longo da cóclea. As frequências características
(CF) (correspondentes ao pico das respostas neuronais das fibras) em função da posição
coclear de onde provêm as fibras, parece ser invariante para muitas espécies animais e
apresenta a seguinte forma empírica [Liberman, 82], [Greenwood, 90]:
( )( )ω π αCF
x Lx A B( ) = −−2 10 1 . (2.40)
Para a espécie humana, as constantes são A≈160, α ≈2,1 e B≈0,8. É evidente que a
sintonização das fibras no nervo coclear terá de estar necessariamente relacionada com as
respostas mecânicas da partição coclear, embora esta relação não esteja completamente
esclarecida. Alguns modelos cocleares que serão abordados seguidamente, definem
explicitamente esta relação. A expressão de ωCF(x) é aproximadamente exponencial excepto
para posições apicais. Na figura 2.6 apresentam-se as curvas correspondentes às equações
(2.39) e (2.40) onde os parâmetros ωc0 e a são ajustados de forma a que as curvas
coincidam para posições basais.
Continuando a descrição da impedância da partição, se a resistência R(x) variar também
exponencialmente, mas com metade do expoente de S(x), isto é, se for proporcional a ω0(x),
significa que o factor de amortecimento associado a esta impedância de 2ª ordem virá
constante. Seja ele δ/2. A impedância da partição apresenta então a seguinte forma:
( )Z xM
ss s x xp ( ) ( ) ( )= + +2
0 02δω ω . (2.41)
Esta definição da impedância reproduz uma propriedade de simetria de escala que se sabe
existir, pelo menos de uma forma aproximada e para posições não muito distantes da base,
nas respostas mecânicas cocleares: para posições x fixas, as respostas em função da
frequência são essencialmente idênticas sofrendo apenas uma translação em função da
frequência de ressonância ω0(x). Analogamente, para frequências de excitação fixas, as
respostas em função de x são também idênticas, deslocadas entre si apenas em função da
frequência de excitação. As curvas de sintonia neuronal apresentam igualmente esta
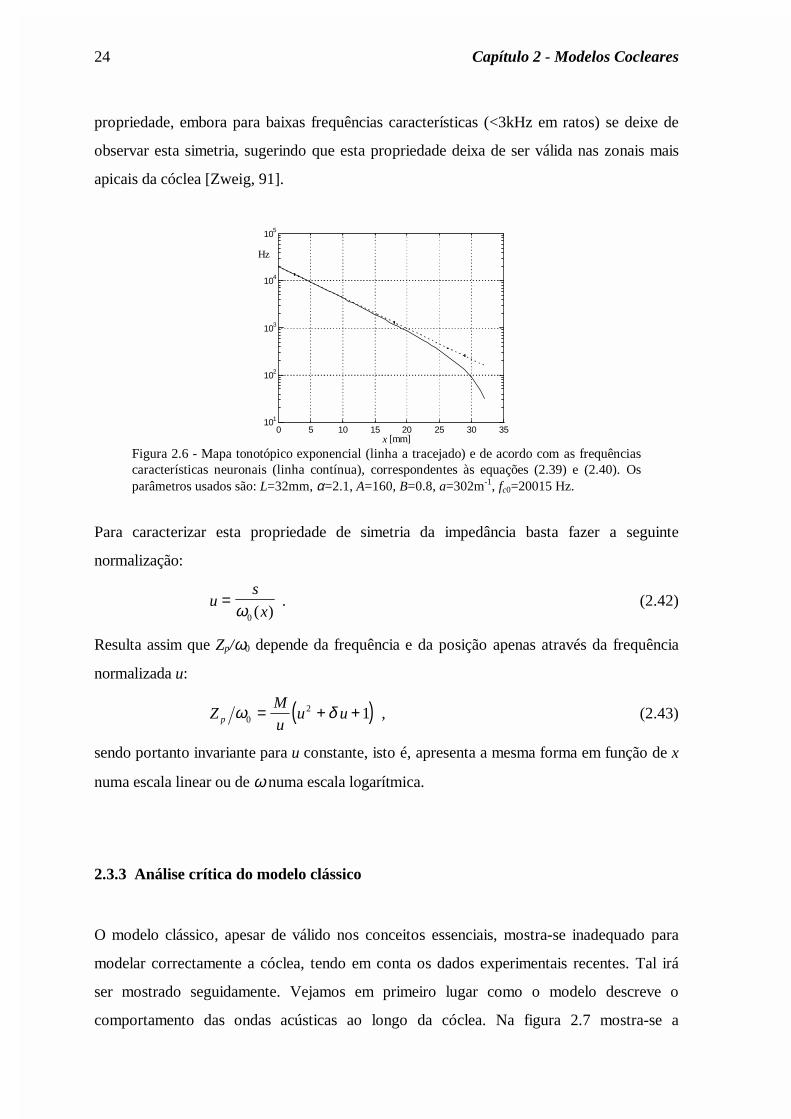
24 Capítulo 2 - Modelos Cocleares
propriedade, embora para baixas frequências características (<3kHz em ratos) se deixe de
observar esta simetria, sugerindo que esta propriedade deixa de ser válida nas zonais mais
apicais da cóclea [Zweig, 91].
0 5 10 15 20 25 30 35101
102
103
104
105
x [mm]
Hz
Figura 2.6 - Mapa tonotópico exponencial (linha a tracejado) e de acordo com as frequênciascaracterísticas neuronais (linha contínua), correspondentes às equações (2.39) e (2.40). Osparâmetros usados são: L=32mm, α=2.1, A=160, B=0.8, a=302m-1, fc0=20015 Hz.
Para caracterizar esta propriedade de simetria da impedância basta fazer a seguinte
normalização:
us
x=
ω0 ( ) . (2.42)
Resulta assim que Zp/ω0 depende da frequência e da posição apenas através da frequência
normalizada u:
( )ZM
uu up ω δ0
2 1= + + , (2.43)
sendo portanto invariante para u constante, isto é, apresenta a mesma forma em função de x
numa escala linear ou de ω numa escala logarítmica.
2.3.3 Análise crítica do modelo clássico
O modelo clássico, apesar de válido nos conceitos essenciais, mostra-se inadequado para
modelar correctamente a cóclea, tendo em conta os dados experimentais recentes. Tal irá
ser mostrado seguidamente. Vejamos em primeiro lugar como o modelo descreve o
comportamento das ondas acústicas ao longo da cóclea. Na figura 2.7 mostra-se a
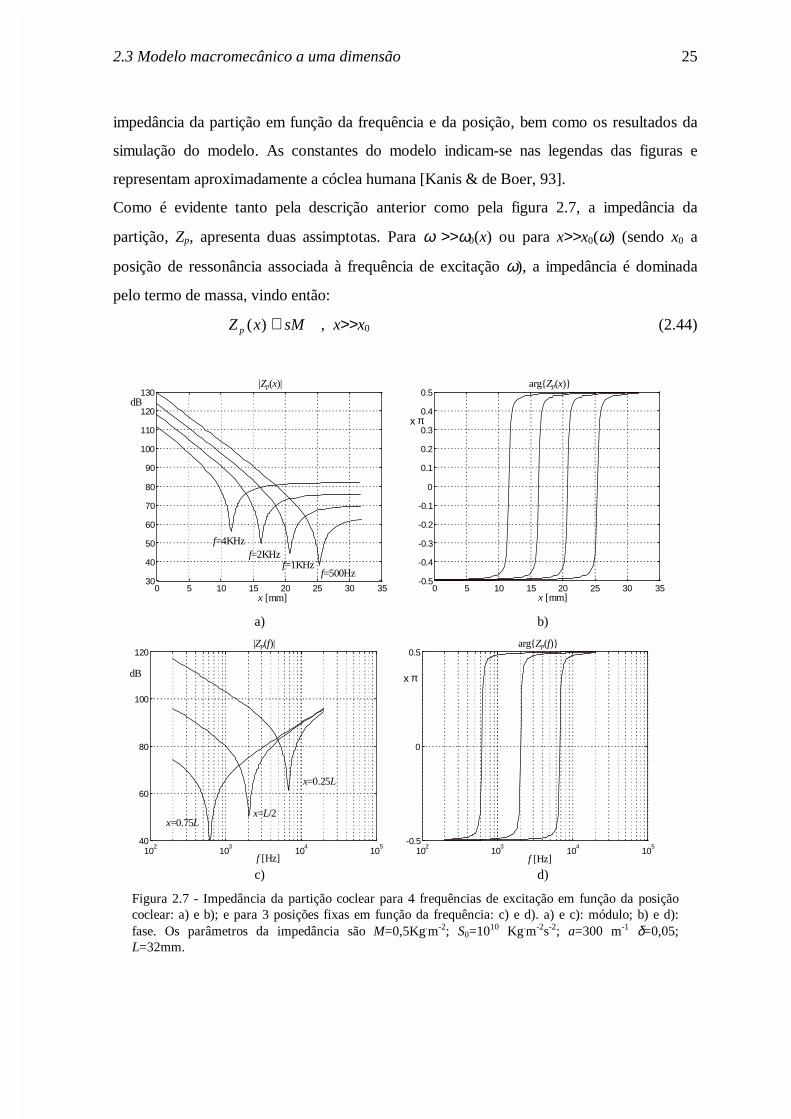
2.3 Modelo macromecânico a uma dimensão 25
impedância da partição em função da frequência e da posição, bem como os resultados da
simulação do modelo. As constantes do modelo indicam-se nas legendas das figuras e
representam aproximadamente a cóclea humana [Kanis & de Boer, 93].
Como é evidente tanto pela descrição anterior como pela figura 2.7, a impedância da
partição, Zp, apresenta duas assimptotas. Para ω >>ω0(x) ou para x>>x0(ω) (sendo x0 a
posição de ressonância associada à frequência de excitação ω), a impedância é dominada
pelo termo de massa, vindo então:
Z x sMp ( ) ≅ , x>>x0 (2.44)
0 5 10 15 20 25 30 35
30
40
50
60
70
80
90
100
110
120
130
x [mm]
dB
|Zp(x)|
f=4KHz f=2KHz
f=1KHz f=500Hz
0 5 10 15 20 25 30 35
-0.5
-0.4
-0.3
-0.2
-0.1
0
0.1
0.2
0.3
0.4
0.5
x [mm]
x π
argZp(x)
a) b)
10
210
310
410
540
60
80
100
120
f [Hz]
dB
|Zp(f)|
x=L/2 x=0.75L
x=0.25L
10
210
310
410
5-0.5
0
0.5
f [Hz]
x π
argZp(f)
c) d)
Figura 2.7 - Impedância da partição coclear para 4 frequências de excitação em função da posiçãococlear: a) e b); e para 3 posições fixas em função da frequência: c) e d). a) e c): módulo; b) e d):fase. Os parâmetros da impedância são M=0,5Kg.m-2; S0=1010 Kg.m-2s-2; a=300 m-1 δ=0,05;L=32mm.
26 Capítulo 2 - Modelos Cocleares
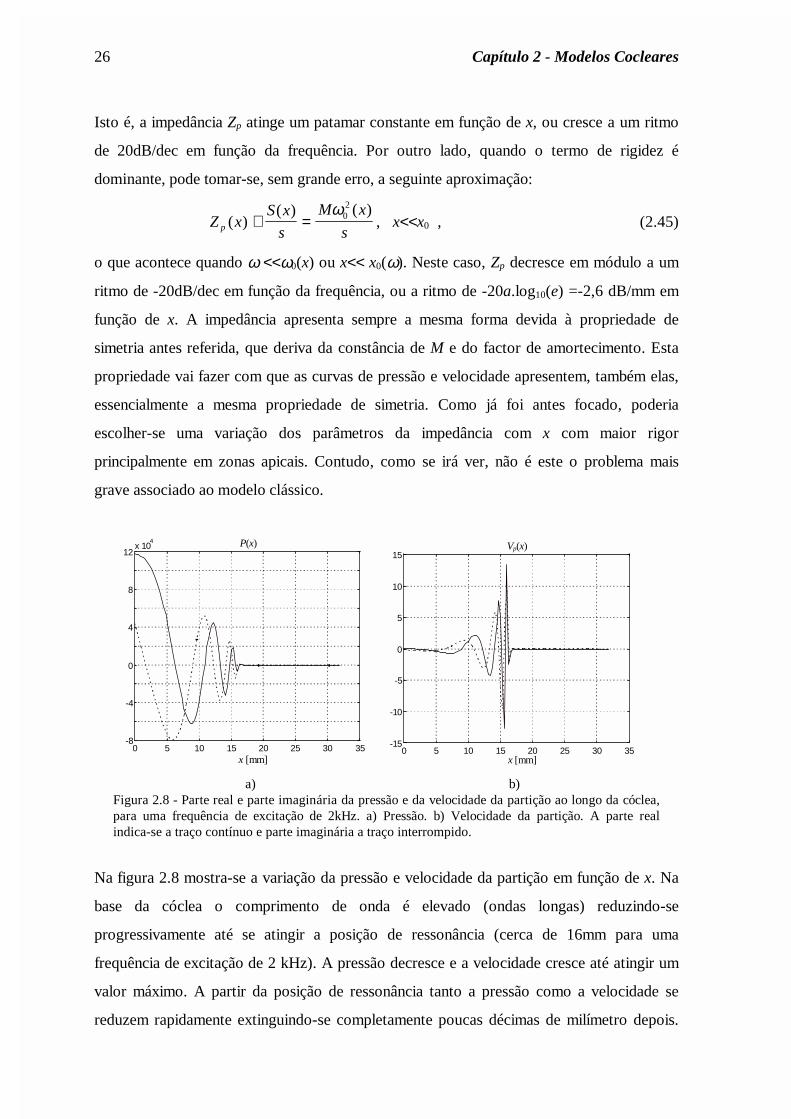
Isto é, a impedância Zp atinge um patamar constante em função de x, ou cresce a um ritmo
de 20dB/dec em função da frequência. Por outro lado, quando o termo de rigidez é
dominante, pode tomar-se, sem grande erro, a seguinte aproximação:
Z xS x
s
M x
sp ( )( ) ( )
≅ =ω0
2
, x<<x0 , (2.45)
o que acontece quando ω <<ω0(x) ou x<< x0(ω). Neste caso, Zp decresce em módulo a um
ritmo de -20dB/dec em função da frequência, ou a ritmo de -20a.log10(e) =-2,6 dB/mm em
função de x. A impedância apresenta sempre a mesma forma devida à propriedade de
simetria antes referida, que deriva da constância de M e do factor de amortecimento. Esta
propriedade vai fazer com que as curvas de pressão e velocidade apresentem, também elas,
essencialmente a mesma propriedade de simetria. Como já foi antes focado, poderia
escolher-se uma variação dos parâmetros da impedância com x com maior rigor
principalmente em zonas apicais. Contudo, como se irá ver, não é este o problema mais
grave associado ao modelo clássico.
0 5 10 15 20 25 30 35
-8
-4
0
4
8
12x 10
4
x [mm]
P(x)
0 5 10 15 20 25 30 35
-15
-10
-5
0
5
10
15
x [mm]
Vp(x)
a) b)Figura 2.8 - Parte real e parte imaginária da pressão e da velocidade da partição ao longo da cóclea,para uma frequência de excitação de 2kHz. a) Pressão. b) Velocidade da partição. A parte realindica-se a traço contínuo e parte imaginária a traço interrompido.
Na figura 2.8 mostra-se a variação da pressão e velocidade da partição em função de x. Na
base da cóclea o comprimento de onda é elevado (ondas longas) reduzindo-se
progressivamente até se atingir a posição de ressonância (cerca de 16mm para uma
frequência de excitação de 2 kHz). A pressão decresce e a velocidade cresce até atingir um
valor máximo. A partir da posição de ressonância tanto a pressão como a velocidade se
reduzem rapidamente extinguindo-se completamente poucas décimas de milímetro depois.
2.3 Modelo macromecânico a uma dimensão 27
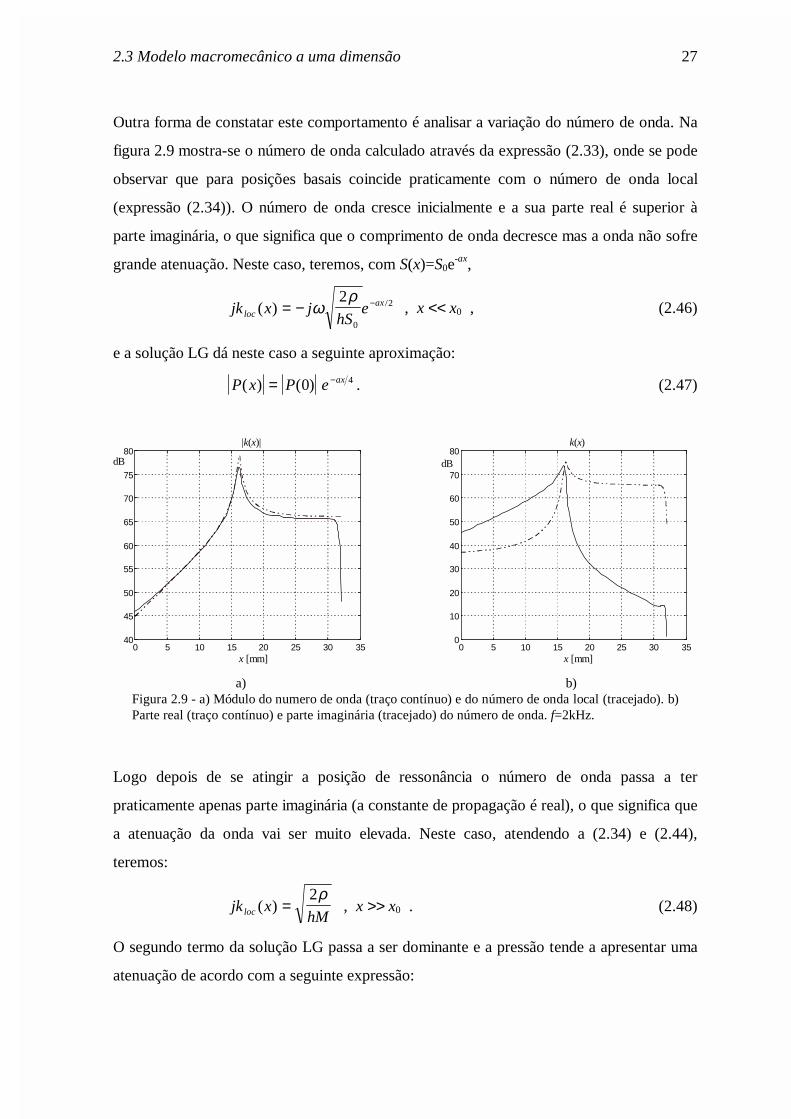
Outra forma de constatar este comportamento é analisar a variação do número de onda. Na
figura 2.9 mostra-se o número de onda calculado através da expressão (2.33), onde se pode
observar que para posições basais coincide praticamente com o número de onda local
(expressão (2.34)). O número de onda cresce inicialmente e a sua parte real é superior à
parte imaginária, o que significa que o comprimento de onda decresce mas a onda não sofre
grande atenuação. Neste caso, teremos, com S(x)=S0e-ax,
jk x jhS
elocax( ) /= − −ω
ρ2
0
2 , x << x0 , (2.46)
e a solução LG dá neste caso a seguinte aproximação:
P x P e ax( ) ( )= −0 4 . (2.47)
0 5 10 15 20 25 30 3540
45
50
55
60
65
70
75
80
x [mm]
dB
|k(x)|
0 5 10 15 20 25 30 350
10
20
30
40
50
60
70
80
x [mm]
dB
k(x)
a) b)Figura 2.9 - a) Módulo do numero de onda (traço contínuo) e do número de onda local (tracejado). b)Parte real (traço contínuo) e parte imaginária (tracejado) do número de onda. f=2kHz.
Logo depois de se atingir a posição de ressonância o número de onda passa a ter
praticamente apenas parte imaginária (a constante de propagação é real), o que significa que
a atenuação da onda vai ser muito elevada. Neste caso, atendendo a (2.34) e (2.44),
teremos:
jk xhMloc ( ) =2ρ
, x >> x0 . (2.48)
O segundo termo da solução LG passa a ser dominante e a pressão tende a apresentar uma
atenuação de acordo com a seguinte expressão:
28 Capítulo 2 - Modelos Cocleares
P xhM
x( ) exp∝ −
2ρ. (2.49e)
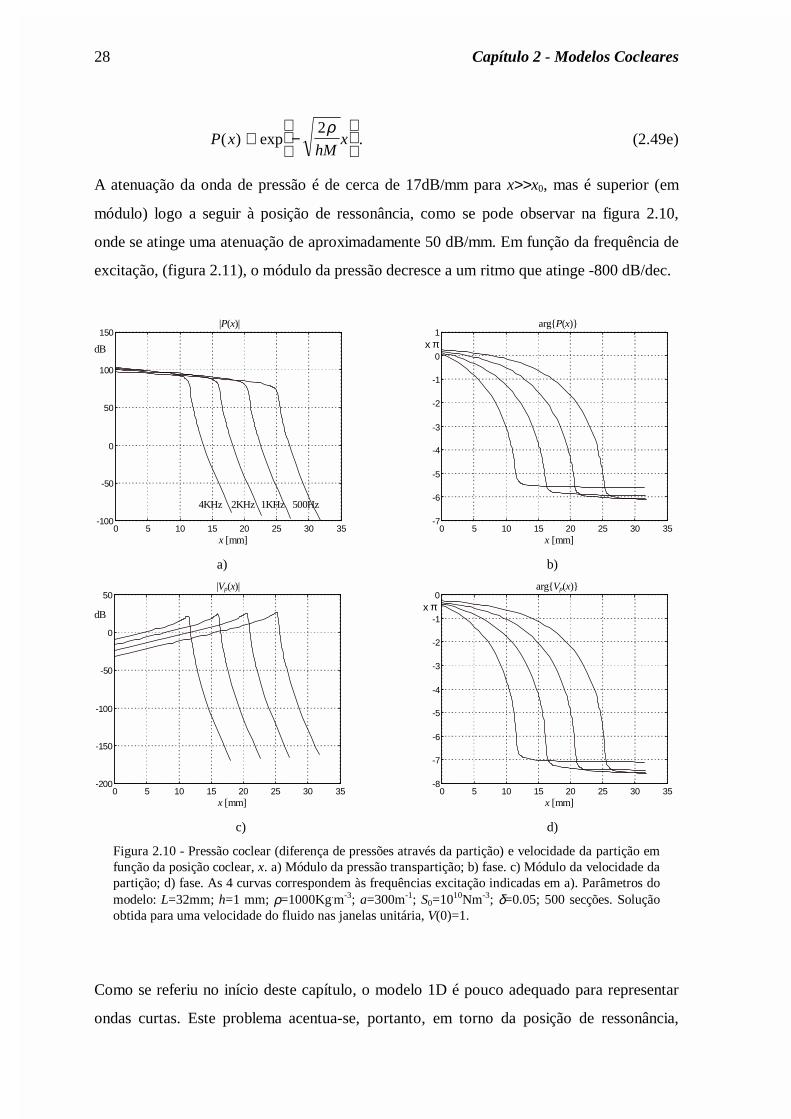
A atenuação da onda de pressão é de cerca de 17dB/mm para x>>x0, mas é superior (em
módulo) logo a seguir à posição de ressonância, como se pode observar na figura 2.10,
onde se atinge uma atenuação de aproximadamente 50 dB/mm. Em função da frequência de
excitação, (figura 2.11), o módulo da pressão decresce a um ritmo que atinge -800 dB/dec.
0 5 10 15 20 25 30 35-100
-50
0
50
100
150
x [mm]
dB
|P(x)|
4KHz 2KHz 1KHz 500Hz
0 5 10 15 20 25 30 35-7
-6
-5
-4
-3
-2
-1
0
1
x [mm]
x π
argP(x)
a) b)
0 5 10 15 20 25 30 35-200
-150
-100
-50
0
50
x [mm]
dB
|Vp(x)|
0 5 10 15 20 25 30 35-8
-7
-6
-5
-4
-3
-2
-1
0
x [mm]
x π
argVp(x)
c) d)
Figura 2.10 - Pressão coclear (diferença de pressões através da partição) e velocidade da partição emfunção da posição coclear, x. a) Módulo da pressão transpartição; b) fase. c) Módulo da velocidade dapartição; d) fase. As 4 curvas correspondem às frequências excitação indicadas em a). Parâmetros domodelo: L=32mm; h=1 mm; ρ=1000Kg.m-3; a=300m-1; S0=1010Nm-3; δ=0.05; 500 secções. Soluçãoobtida para uma velocidade do fluido nas janelas unitária, V(0)=1.
Como se referiu no início deste capítulo, o modelo 1D é pouco adequado para representar
ondas curtas. Este problema acentua-se, portanto, em torno da posição de ressonância,
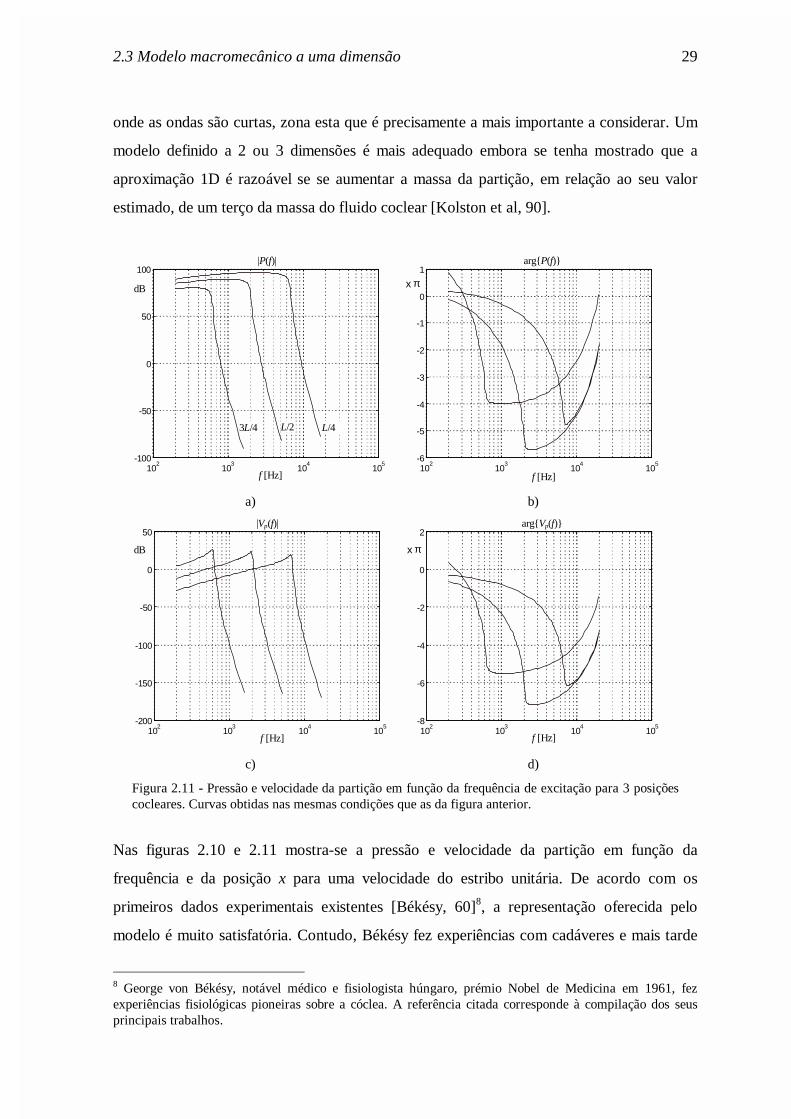
2.3 Modelo macromecânico a uma dimensão 29
onde as ondas são curtas, zona esta que é precisamente a mais importante a considerar. Um
modelo definido a 2 ou 3 dimensões é mais adequado embora se tenha mostrado que a
aproximação 1D é razoável se se aumentar a massa da partição, em relação ao seu valor
estimado, de um terço da massa do fluido coclear [Kolston et al, 90].
10
210
310
410
5-100
-50
0
50
100
f [Hz]
dB
|P(f)|
3L/4 L/4L/2
10
210
310
410
5-6
-5
-4
-3
-2
-1
0
1
f [Hz]
x π
argP(f)
a) b)
10
210
310
410
5-200
-150
-100
-50
0
50
f [Hz]
dB
|Vp(f)|
10
210
310
410
5-8
-6
-4
-2
0
2
f [Hz]
x π
argVp(f)
c) d)
Figura 2.11 - Pressão e velocidade da partição em função da frequência de excitação para 3 posiçõescocleares. Curvas obtidas nas mesmas condições que as da figura anterior.
Nas figuras 2.10 e 2.11 mostra-se a pressão e velocidade da partição em função da
frequência e da posição x para uma velocidade do estribo unitária. De acordo com os
primeiros dados experimentais existentes [Békésy, 60]8, a representação oferecida pelo
modelo é muito satisfatória. Contudo, Békésy fez experiências com cadáveres e mais tarde
8 George von Békésy, notável médico e fisiologista húngaro, prémio Nobel de Medicina em 1961, fezexperiências fisiológicas pioneiras sobre a cóclea. A referência citada corresponde à compilação dos seusprincipais trabalhos.
30 Capítulo 2 - Modelos Cocleares
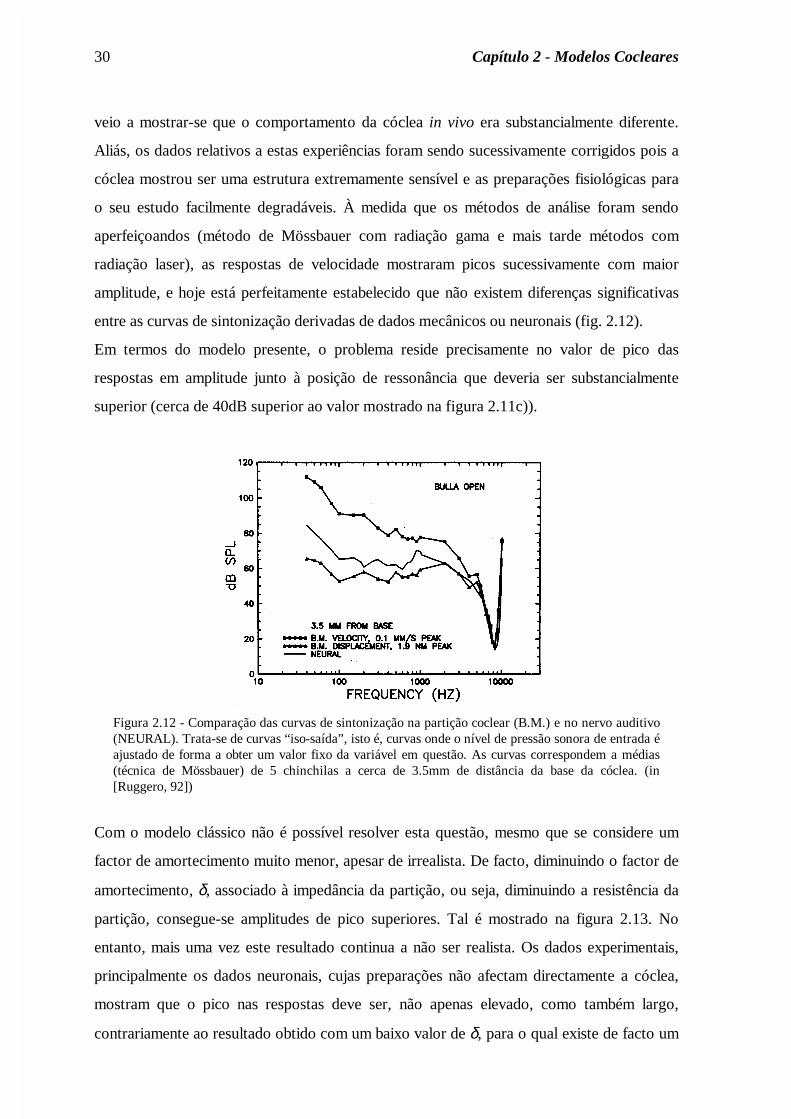
veio a mostrar-se que o comportamento da cóclea in vivo era substancialmente diferente.
Aliás, os dados relativos a estas experiências foram sendo sucessivamente corrigidos pois a
cóclea mostrou ser uma estrutura extremamente sensível e as preparações fisiológicas para
o seu estudo facilmente degradáveis. À medida que os métodos de análise foram sendo
aperfeiçoandos (método de Mössbauer com radiação gama e mais tarde métodos com
radiação laser), as respostas de velocidade mostraram picos sucessivamente com maior
amplitude, e hoje está perfeitamente estabelecido que não existem diferenças significativas
entre as curvas de sintonização derivadas de dados mecânicos ou neuronais (fig. 2.12).
Em termos do modelo presente, o problema reside precisamente no valor de pico das
respostas em amplitude junto à posição de ressonância que deveria ser substancialmente
superior (cerca de 40dB superior ao valor mostrado na figura 2.11c)).
Figura 2.12 - Comparação das curvas de sintonização na partição coclear (B.M.) e no nervo auditivo(NEURAL). Trata-se de curvas “iso-saída”, isto é, curvas onde o nível de pressão sonora de entrada éajustado de forma a obter um valor fixo da variável em questão. As curvas correspondem a médias(técnica de Mössbauer) de 5 chinchilas a cerca de 3.5mm de distância da base da cóclea. (in[Ruggero, 92])
Com o modelo clássico não é possível resolver esta questão, mesmo que se considere um
factor de amortecimento muito menor, apesar de irrealista. De facto, diminuindo o factor de
amortecimento, δ, associado à impedância da partição, ou seja, diminuindo a resistência da
partição, consegue-se amplitudes de pico superiores. Tal é mostrado na figura 2.13. No
entanto, mais uma vez este resultado continua a não ser realista. Os dados experimentais,
principalmente os dados neuronais, cujas preparações não afectam directamente a cóclea,
mostram que o pico nas respostas deve ser, não apenas elevado, como também largo,
contrariamente ao resultado obtido com um baixo valor de δ, para o qual existe de facto um
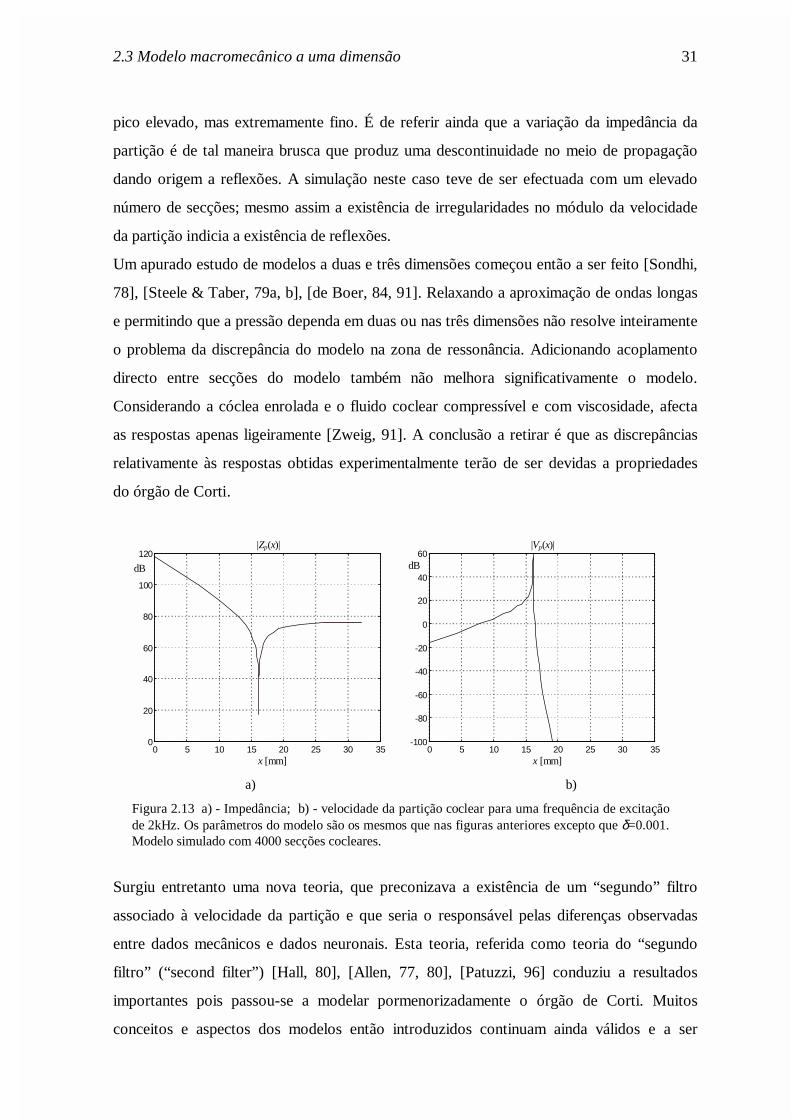
2.3 Modelo macromecânico a uma dimensão 31
pico elevado, mas extremamente fino. É de referir ainda que a variação da impedância da
partição é de tal maneira brusca que produz uma descontinuidade no meio de propagação
dando origem a reflexões. A simulação neste caso teve de ser efectuada com um elevado
número de secções; mesmo assim a existência de irregularidades no módulo da velocidade
da partição indicia a existência de reflexões.
Um apurado estudo de modelos a duas e três dimensões começou então a ser feito [Sondhi,
78], [Steele & Taber, 79a, b], [de Boer, 84, 91]. Relaxando a aproximação de ondas longas
e permitindo que a pressão dependa em duas ou nas três dimensões não resolve inteiramente
o problema da discrepância do modelo na zona de ressonância. Adicionando acoplamento
directo entre secções do modelo também não melhora significativamente o modelo.
Considerando a cóclea enrolada e o fluido coclear compressível e com viscosidade, afecta
as respostas apenas ligeiramente [Zweig, 91]. A conclusão a retirar é que as discrepâncias
relativamente às respostas obtidas experimentalmente terão de ser devidas a propriedades
do órgão de Corti.
0 5 10 15 20 25 30 35
0
20
40
60
80
100
120|Zp(x)|
x [mm]
dB
0 5 10 15 20 25 30 35
-100
-80
-60
-40
-20
0
20
40
60|Vp(x)|
x [mm]
dB
a) b)
Figura 2.13 a) - Impedância; b) - velocidade da partição coclear para uma frequência de excitaçãode 2kHz. Os parâmetros do modelo são os mesmos que nas figuras anteriores excepto que δ=0.001.Modelo simulado com 4000 secções cocleares.
Surgiu entretanto uma nova teoria, que preconizava a existência de um “segundo” filtro
associado à velocidade da partição e que seria o responsável pelas diferenças observadas
entre dados mecânicos e dados neuronais. Esta teoria, referida como teoria do “segundo
filtro” (“second filter”) [Hall, 80], [Allen, 77, 80], [Patuzzi, 96] conduziu a resultados
importantes pois passou-se a modelar pormenorizadamente o órgão de Corti. Muitos
conceitos e aspectos dos modelos então introduzidos continuam ainda válidos e a ser

32 Capítulo 2 - Modelos Cocleares
utilizados. No entanto, a teoria em si, viria a ser desvalorizada dada a concordância que se
veio a mostrar posteriormente entre os dados mecânicos e neuronais [Robles et al, 86] (ver
figura 2.12).
A descoberta das emissões otoacústicas espontâneas veio levantar um novo problema na
modelação coclear: como poderia um sistema passivo e aparentemente estável, produzir
espontaneamente sons para o exterior? A hipótese de a cóclea poder ser um sistema activo
começou a ganhar forma tendo sido inicialmente proposta por Davis [Davis, 83] e Kim,
[Kim, 84] e vindo a solidificar-se com a descoberta da mobilidade das células ciliadas
externas. Contudo, a teoria da actividade coclear, conhecida também como teoria da
amplificação coclear, gerou uma enorme controvérsia que não está, ainda hoje,
inteiramente resolvida [de Boer, 83], [Allen, 85], [Kolston et al, 90], [Allen & Neely, 1992].
Em conclusão, o modelo clássico é válido nos seus pressupostos macromecânicos
fundamentais, relativos à física das ondas progressivas na cóclea (“traveling waves”). As
simplificações quanto à dimensão do modelo bem como à sua geometria parecem ser
aceitáveis desde que se defina com maior rigor e pormenor a impedância da partição
incluindo nesta o efeito das principais estruturas do órgão de Corti. Este assunto é
abordado na secção seguinte.
Antes de abordar os modelos micromecânicos é conveniente referir dois outros aspectos
importantes na modelação coclear. O primeiro destes aspectos diz respeito à impedância de
entrada da cóclea e o segundo à modelação do ouvido externo e médio.
2.4 Impedância de entrada da cóclea
Existem muitos estudos experimentais sobre a medida da impedância de entrada da cóclea
(por exemplo [Lynch et al, 82], [Nedzelnitsky, 80], [Rabinowitz, 81], [Rosowski, 96]),
sendo a sua caracterização essencial no estudo do ouvido médio. Por outro lado, a análise
desta impedância em modelos cocleares constitui um aspecto importante a considerar para a
verificação da concordância dos modelos com dados conhecidos.
No modelo coclear anteriormente definido, a impedância de entrada da cóclea corresponde
à impedância Zr(x) para x=0. É a relação entre a diferença de pressões junto das janelas e a
velocidade do fluido na mesma localização que é igual à velocidade do estribo.

2.4 Impedância de entrada da cóclea 33
Junto às janelas a impedância da partição é dominada pela sua rigidez, a não ser para
frequências de excitação próximas da frequência de ressonância da base da cóclea. Assim,
uma análise assimptótica do modelo é válida o que permite determinar com facilidade a
impedância de entrada da cóclea. Vejamos qual a sua expressão para o modelo clássico.
Para posições cocleares inferiores à posição de ressonância, x→0, (e com ω<<ω0), teremos,
tal como em (2.45):
Z x S e jpax( ) ≅ −
0 ω . (2.50)
Nesta situação, com x=0, tanto o número de onda local como a impedância característica
local são reais (ver (2.14)):
k j Z ZhS
Dloc ser par( ) ( )0 02
00= − = =ω
ρ , (2.51)
Z Z Z hS Rloc ser par in0 00 0 2( ) ( )= = =ρ . (2.52)
Como iremos ver, Rin é o valor assimptótico da impedância de entrada, Zin, mas apenas
correcto para frequências intermédias. Podemos saber o valor da impedância de entrada
com a aproximação (2.50), pois nesta situação o modelo tem uma solução analítica. A
equação do modelo, (2.8), passa a ser a seguinte:
d P x
dxD e P xax
2
2 02 0
( )( )+ = . (2.53)
Fazendo a seguinte mudança de variáveis:
vD
ae
k
k
k
kxax loc
loc
loc
loc
= =′
′
2 0
0
0
00 2
2/ ( )
( )exp
( )
( ) , (2.54)
esta equação diferencial transforma-se numa equação de Bessel. A solução é, [de Boer, 80],
[Koshigoe et al, 83]:
P v cH v( ) ( )( )= 02 , (2.55)
onde H02( ) é a função cilíndrica de Hankel de ordem zero, 2ª espécie [Kinsler et al, 82] 9,
onde c é uma constante. Atendendo à definição de Zr(x) (ver expressão (2.30)), facilmente
se pode verificar que a impedância de entrada resulta na seguinte expressão:
9 H x J x jY xn n n
( ) ( ) ( ) ( )2 = − onde Jn(x) e Yn(x) são as funções de Bessel de ordem n de 1ª e 2ª espécie,
respectivamente. dH x dx H x02
12( ) ( )( ) ( )= − . Para x>>2π, H x
xen
j xn
( ) ( )2 2 42→
− − −
π
π π
.
34 Capítulo 2 - Modelos Cocleares
Z jRH v
H vin in= 02
0
12
0
( )
( )
( )
( ) . (2.56)
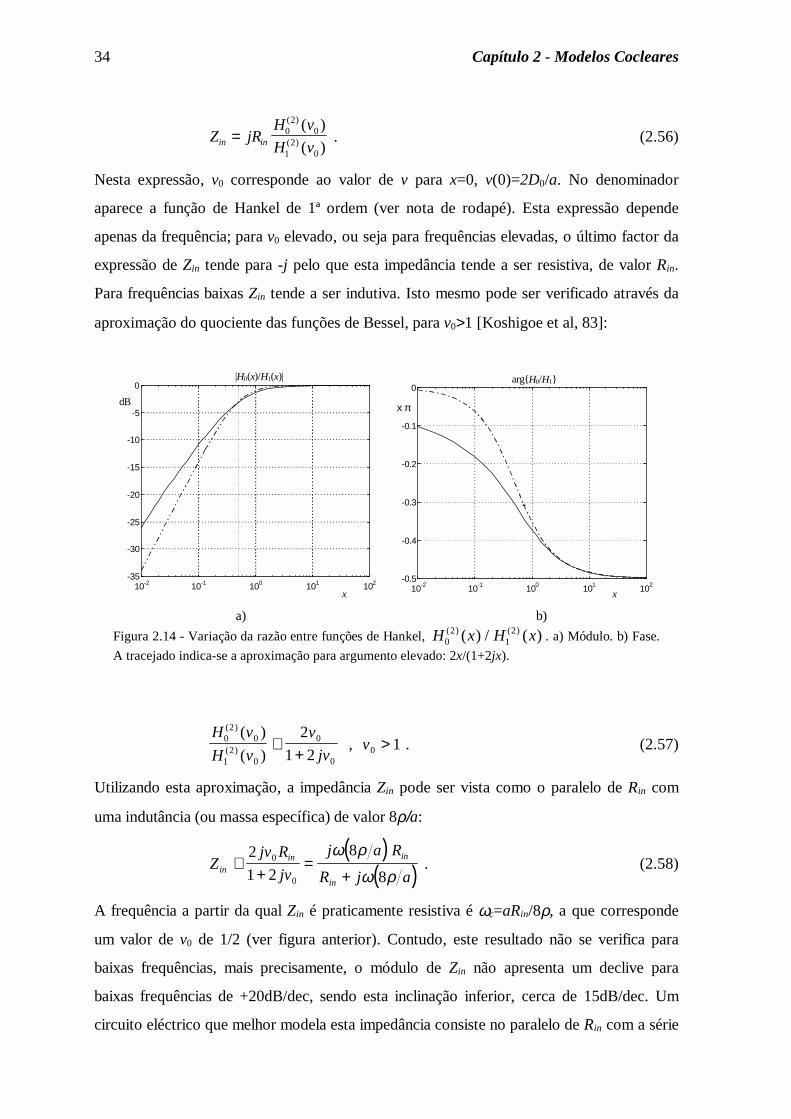
Nesta expressão, v0 corresponde ao valor de v para x=0, v(0)=2D0/a. No denominador
aparece a função de Hankel de 1ª ordem (ver nota de rodapé). Esta expressão depende
apenas da frequência; para v0 elevado, ou seja para frequências elevadas, o último factor da
expressão de Zin tende para -j pelo que esta impedância tende a ser resistiva, de valor Rin.
Para frequências baixas Zin tende a ser indutiva. Isto mesmo pode ser verificado através da
aproximação do quociente das funções de Bessel, para v0>1 [Koshigoe et al, 83]:
10-2 10-1 100 101 102-35
-30
-25
-20
-15
-10
-5
0|H0(x)/H1(x)|
x
dB
10-2 10-1 100 101 102-0.5
-0.4
-0.3
-0.2
-0.1
0argH0/H1
x
x π
a) b)
Figura 2.14 - Variação da razão entre funções de Hankel, H x H x02
12( ) ( )( ) / ( ) . a) Módulo. b) Fase.
A tracejado indica-se a aproximação para argumento elevado: 2x/(1+2jx).
H v
H v
v
jvv0
20
12
0
0
00
2
1 21
( )
( )
( )
( )≅
+> , . (2.57)
Utilizando esta aproximação, a impedância Zin pode ser vista como o paralelo de Rin com
uma indutância (ou massa específica) de valor 8ρ/a:
( )( )Z
jv R
jv
j a R
R j ain
in in
in
≅+
=+
2
1 2
8
80
0
ω ρ
ω ρ
. (2.58)
A frequência a partir da qual Zin é praticamente resistiva é ωc=aRin/8ρ, a que corresponde
um valor de v0 de 1/2 (ver figura anterior). Contudo, este resultado não se verifica para
baixas frequências, mais precisamente, o módulo de Zin não apresenta um declive para
baixas frequências de +20dB/dec, sendo esta inclinação inferior, cerca de 15dB/dec. Um
circuito eléctrico que melhor modela esta impedância consiste no paralelo de Rin com a série

2.4 Impedância de entrada da cóclea 35
de uma resistência e uma indutância. É este o modelo inferido por Lynch [Lynch et al, 82] a
partir de resultados experimentais em gatos.
É interessante fazer aqui um parênteses para verificar que a solução WKB (ou LG)
corresponde precisamente ao valor assimptótico de P(v). De facto, para v>2π, resulta
P v cv
k
k xeloc
loc
ax( )( )
( )→ ∝ = −2 0 4
π . (2.59)
A pressão P(x), normalizada à pressão nas janelas, será portanto:
P x
P
H v
H ve ax( )
( )
( )
( )
( )
( )00
2
02
0
4= → − , (2.60)
onde v0 corresponde ao valor de v para x=0: v(0)=2D0/a. Isto significa que a onda
estacionária de pressão se atenua numa escala logarítmica (em dB), a um ritmo de 5a/ln(10)
ou cerca de 0,65dB/mm, antes de atingir a posição de ressonância. Por outro lado, a
velocidade da partição normalizada à velocidade do estribo, V(0), será:
V x
V
P x
P
Z
Z x
j R
Se
H v
H vp in
p
in ax( )
( )
( )
( ) ( )
( )
( )
( )
( )0 0 0
02
12
0
= − =− ω
, (2.61)
e portanto, esta velocidade normalizada, em módulo, tende para ser proporcional à
frequência e exponencial com expoente 3ax/4, isto é, cresce cerca de 2dB/mm (ver fig.
2.10c).
Voltando de novo à análise da impedância de entrada da cóclea, na figura 2.15 apresenta-se
a impedância Zin acústica, Zin(a) = Zin/A(0), calculada de acordo com a expressão (2.56),
conjuntamente com a impedância acústica de entrada calculada numericamente. Observa-se
que, de facto, esta solução assimptótica é correcta excepto para frequências próximas da
frequência de ressonância correspondente à base da cóclea. Tal seria de esperar, pois a
aproximação (2.50) é válida apenas para ω<<ω0(x). O declive do módulo da impedância
para baixas frequências é inferior a 20dB/dec (cerca de 15dB/dec), resultado este
condizente com os dados obtidos experimentalmente (fig. 2.16). No entanto, devido ao
mapa tonotópico exponencial, a frequência de ressonância no helicotrema é cerca de
180Hz; para frequências de excitação menores, a onda propaga-se até ao helicotrema, onde
chega ainda pouco atenuada, sendo portanto reflectida por desadaptação. Daí a oscilação
no módulo de Zin para frequências inferiores a cerca de 200Hz. Com um factor de
amortecimento menor as reflexões são, obviamente, de maior amplitude. Com a impedância
da partição definida em termos do mapa tonotópico da expressão (2.40), as reflexões são
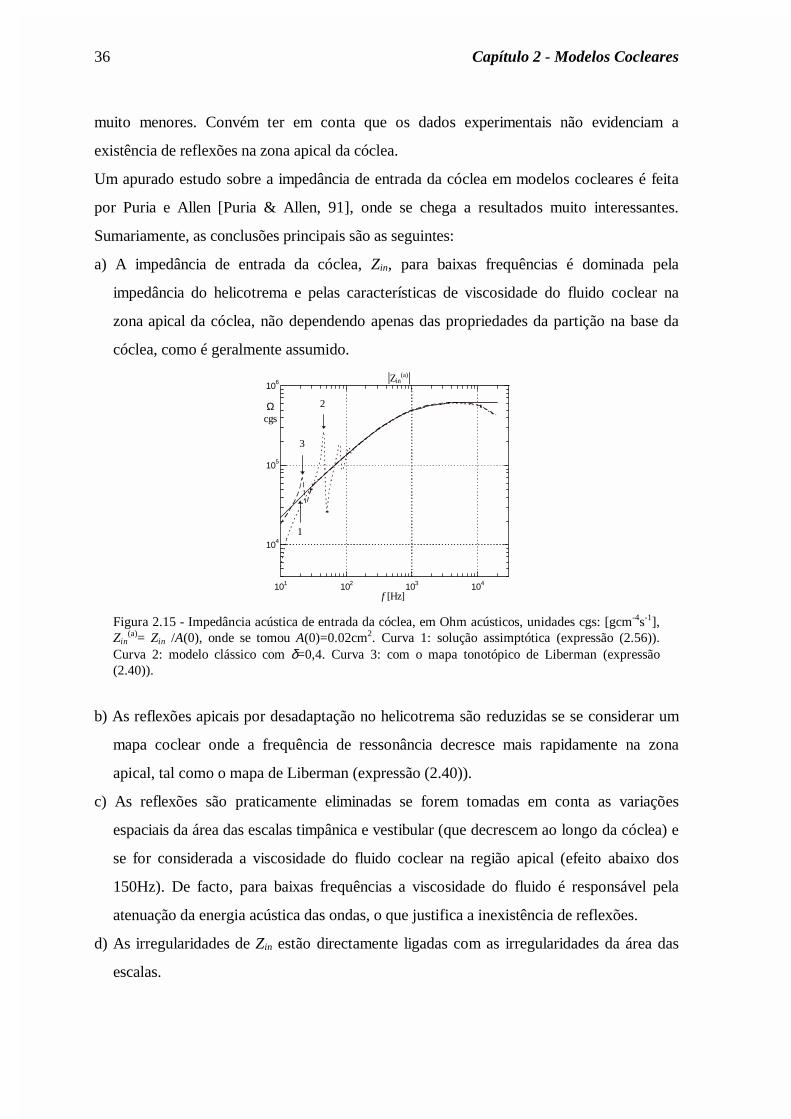
36 Capítulo 2 - Modelos Cocleares
muito menores. Convém ter em conta que os dados experimentais não evidenciam a
existência de reflexões na zona apical da cóclea.
Um apurado estudo sobre a impedância de entrada da cóclea em modelos cocleares é feita
por Puria e Allen [Puria & Allen, 91], onde se chega a resultados muito interessantes.
Sumariamente, as conclusões principais são as seguintes:
a) A impedância de entrada da cóclea, Zin, para baixas frequências é dominada pela
impedância do helicotrema e pelas características de viscosidade do fluido coclear na
zona apical da cóclea, não dependendo apenas das propriedades da partição na base da
cóclea, como é geralmente assumido.
101 102 103 104
104
105
106
f [Hz]
Ωcgs
|Zin(a)|
2
3
1
Figura 2.15 - Impedância acústica de entrada da cóclea, em Ohm acústicos, unidades cgs: [gcm-4s-1],Zin
(a)= Zin /A(0), onde se tomou A(0)=0.02cm2. Curva 1: solução assimptótica (expressão (2.56)).Curva 2: modelo clássico com δ=0,4. Curva 3: com o mapa tonotópico de Liberman (expressão(2.40)).
b) As reflexões apicais por desadaptação no helicotrema são reduzidas se se considerar um
mapa coclear onde a frequência de ressonância decresce mais rapidamente na zona
apical, tal como o mapa de Liberman (expressão (2.40)).
c) As reflexões são praticamente eliminadas se forem tomadas em conta as variações
espaciais da área das escalas timpânica e vestibular (que decrescem ao longo da cóclea) e
se for considerada a viscosidade do fluido coclear na região apical (efeito abaixo dos
150Hz). De facto, para baixas frequências a viscosidade do fluido é responsável pela
atenuação da energia acústica das ondas, o que justifica a inexistência de reflexões.
d) As irregularidades de Zin estão directamente ligadas com as irregularidades da área das
escalas.
2.4 Impedância de entrada da cóclea 37
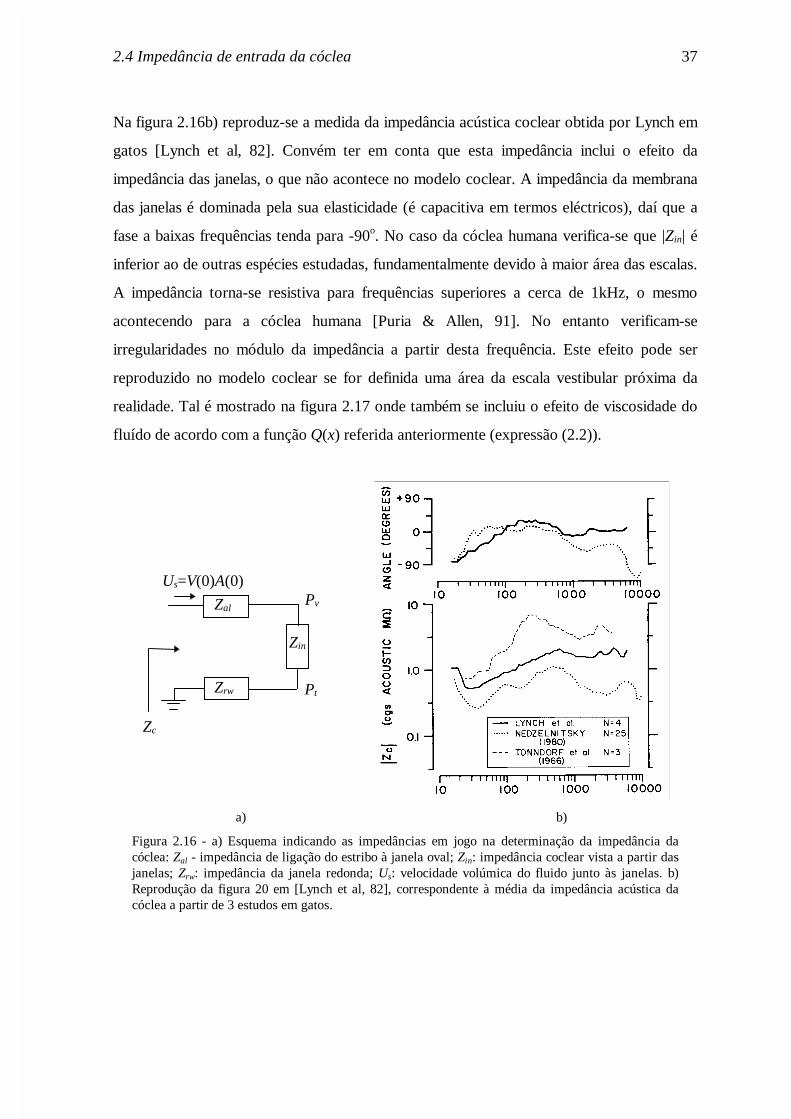
Na figura 2.16b) reproduz-se a medida da impedância acústica coclear obtida por Lynch em
gatos [Lynch et al, 82]. Convém ter em conta que esta impedância inclui o efeito da
impedância das janelas, o que não acontece no modelo coclear. A impedância da membrana
das janelas é dominada pela sua elasticidade (é capacitiva em termos eléctricos), daí que a
fase a baixas frequências tenda para -90o. No caso da cóclea humana verifica-se que |Zin| é
inferior ao de outras espécies estudadas, fundamentalmente devido à maior área das escalas.
A impedância torna-se resistiva para frequências superiores a cerca de 1kHz, o mesmo
acontecendo para a cóclea humana [Puria & Allen, 91]. No entanto verificam-se
irregularidades no módulo da impedância a partir desta frequência. Este efeito pode ser
reproduzido no modelo coclear se for definida uma área da escala vestibular próxima da
realidade. Tal é mostrado na figura 2.17 onde também se incluiu o efeito de viscosidade do
fluído de acordo com a função Q(x) referida anteriormente (expressão (2.2)).
a) b)
Figura 2.16 - a) Esquema indicando as impedâncias em jogo na determinação da impedância dacóclea: Zal - impedância de ligação do estribo à janela oval; Zin: impedância coclear vista a partir dasjanelas; Zrw: impedância da janela redonda; Us: velocidade volúmica do fluido junto às janelas. b)Reprodução da figura 20 em [Lynch et al, 82], correspondente à média da impedância acústica dacóclea a partir de 3 estudos em gatos.
Zal
Zin
Us=V(0)A(0)
Zrw
Pv
Pt
Zc
38 Capítulo 2 - Modelos Cocleares
0 5 10 15 20 25 30 35
10-3
10-2
10-1
x [mm]
cm2
A(x)
10
110
210
310
4
0.1
1
f [Hz]
MΩ|Zin|
2
a) b)
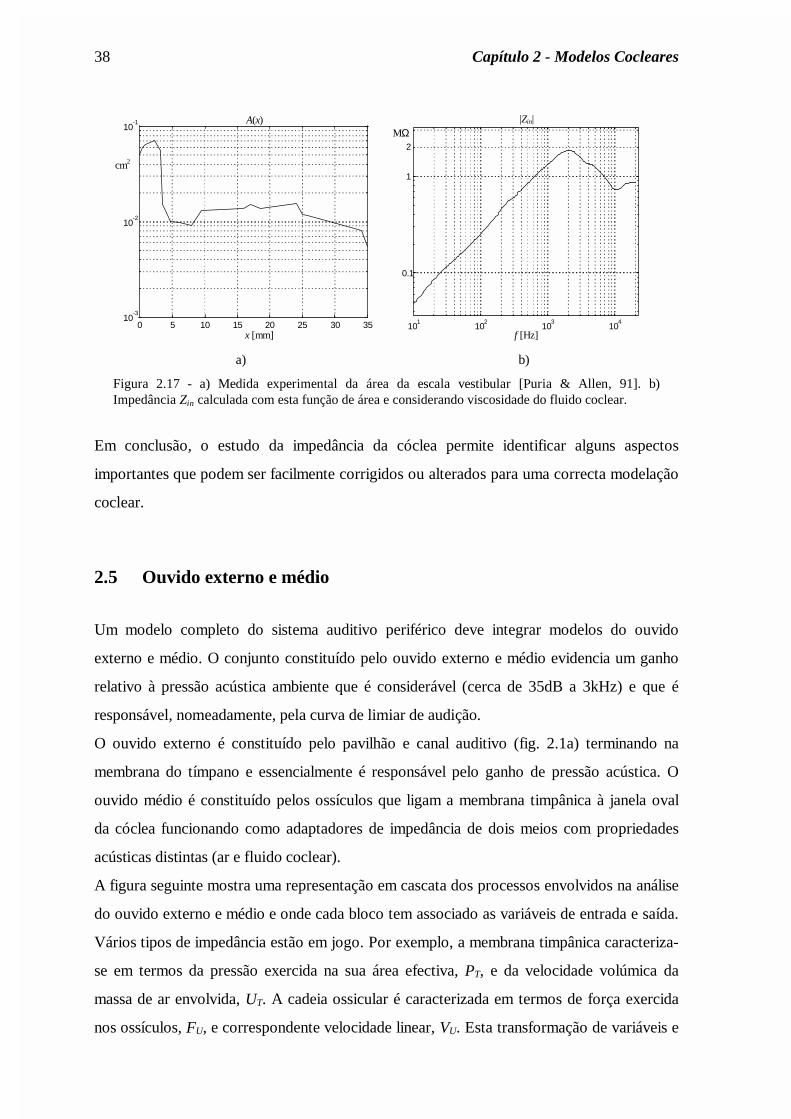
Figura 2.17 - a) Medida experimental da área da escala vestibular [Puria & Allen, 91]. b)Impedância Zin calculada com esta função de área e considerando viscosidade do fluido coclear.
Em conclusão, o estudo da impedância da cóclea permite identificar alguns aspectos
importantes que podem ser facilmente corrigidos ou alterados para uma correcta modelação
coclear.
2.5 Ouvido externo e médio
Um modelo completo do sistema auditivo periférico deve integrar modelos do ouvido
externo e médio. O conjunto constituído pelo ouvido externo e médio evidencia um ganho
relativo à pressão acústica ambiente que é considerável (cerca de 35dB a 3kHz) e que é
responsável, nomeadamente, pela curva de limiar de audição.
O ouvido externo é constituído pelo pavilhão e canal auditivo (fig. 2.1a) terminando na
membrana do tímpano e essencialmente é responsável pelo ganho de pressão acústica. O
ouvido médio é constituído pelos ossículos que ligam a membrana timpânica à janela oval
da cóclea funcionando como adaptadores de impedância de dois meios com propriedades
acústicas distintas (ar e fluido coclear).
A figura seguinte mostra uma representação em cascata dos processos envolvidos na análise
do ouvido externo e médio e onde cada bloco tem associado as variáveis de entrada e saída.
Vários tipos de impedância estão em jogo. Por exemplo, a membrana timpânica caracteriza-
se em termos da pressão exercida na sua área efectiva, PT, e da velocidade volúmica da
massa de ar envolvida, UT. A cadeia ossicular é caracterizada em termos de força exercida
nos ossículos, FU, e correspondente velocidade linear, VU. Esta transformação de variáveis e

2.5 Ouvido externo e médio 39
impedâncias tem como equivalente eléctrico um transformador com razão de transformação
igual à área efectiva da membrana timpânica.
Figura 2.18 - Representação em cascata das funções de transferência que se podem identificar noouvido externo e médio (in [Rosowski, 96]).
Um dos modelos mais pormenorizados do ouvido externo e médio é o modelo desenvolvido
por Rosowski [Rosowski, 96] e que será utilizado neste trabalho com ligeiras modificações.
2.5.1 Ouvido externo
A função de transferência do ouvido externo inclui o efeito da difracção do som pelo corpo,
cabeça e pavilhão auditivo e é altamente dependente da direcção da incidência do som. Para
aplicações de processamento de sinal monoaural um modelo simples que tome em conta
apenas uma direcção é suficiente. Rososwki, no seu modelo considera a cabeça como uma
esfera rígida e calcula o ganho que relaciona a pressão sonora na superfície da esfera rígida
com uma onda plana de pressão de estímulo, PPW. Este ganho resulta numa expressão muito
complicada, definida por uma série com funções de Bessel e polinómios de Legendre. Neste
trabalho considera-se um modelo mais simples que utiliza a aproximação de Bauer [Bauer,
67], em termos de um circuito equivalente representado na figura 2.19. Na caracterização
deste circuito assume-se que o principal obstáculo que a onda incidente encontra pode ser
representado por uma esfera de raio efectivo, r, com uma abertura (pavilhão auditivo) de
área A0. A impedância constituída pelo paralelo da resistência Rr e indutância Mr, representa
a impedância acústica de radiação (em circuito aberto) da abertura correspondente ao
pavilhão auditivo. As equações dos vários elementos deste circuito equivalente são as
seguintes [Giguèrre & Woodland, 94]:

40 Capítulo 2 - Modelos Cocleares
Mr
Rc
rM
AR
c
Asa
sa
ra
ra= = = =
ρπ
ρπ
ρπ
ρ2
0 72
0 0 ,
,
,
., (2.62)
onde ρa é a densidade do ar e c a velocidade do som no ar. Os valores dos parâmetros estão
indicados na legenda da figura 2.19.
As cavidades do pavilhão auditivo e do canal auditivo são modelados, respectivamente, por
uma trompa e por um tubo cujas propriedades acústicas podem ser caracterizadas através
de quadripólos definidos pelos seguintes parâmetros de transmissão [Malecki, 69],
[Rosowski, 96] :
A D kl B jZ kl C j kl ZT T T T T T T= = = =cos( ) , sin( ) , sin( ) / 0 0 . (2.63a)
k c Z c Aa l= =ω ρ , 0 . (2.63b)
A ec e c e
jbB e
j bl
A bHal
jbl jbl
Hal a H
l
H
H H
H=−
=−
2 1
2 ,
sin( )ωρ , (2.63c)
C eA a b bl
j bD e
c e c e
jbHal l H
aH
aljbl jbl
H H
H H
= −+
=−− −
−( ) sin( ),
2 22 1
2ωρ . (2.63d)
b k a c a jb c a jb A A elalH= − = − − = − + =2 2
1 2 02 , , , . (2.63e)
Rs
PPW 2PPW
Rr
AH BH
CH DH
AT BT
CT DT
PEX
+
_
UEX UECUT
PEC PT
+
_
+
_
Difracção do som Pavilhão (trompa) Canal auditivo (tubo)
PPW
lT
lH
A0
e2ax
UEXUTAl
PTPEX
0 x
MsMr
ZiTZECZEX
Figura 2.19 - Modelo do ouvido externo. A relação entre a pressão externa no pavilhão auditivo, PEX, ea pressão da onda plana incidente, PPW, é representada pelo circuito equivalente de Bauer (ver texto). Opavilhão auditivo é representado por uma trompa de secção exponencial, A(x)=A0e
2ax, de comprimentolH. O canal auditivo é representado por um tubo de comprimento lT e de secção Al. Trompa e tubo sãocaracterizados por matrizes de parâmetros de transmissão. Os valores dos parâmetros tomados para estemodelo são os seguintes: r=0.1m; ρa=1.21Kg/m3; c=343m/s; A0=4.4x10-4m2; Al=A0/10; lH=0.01m;lT=0.02m.

2.5 Ouvido externo e médio 41
Esta caracterização pressupõe um meio sem perdas bem como a propagação por ondas
planas (baixas frequências). Para considerar perdas basta tomar o número de onda k1 com
um factor de atenuação α : k1=k−jα [Kinsler el al, 83]. Os valores dos parâmetros são
indicados na legenda da figura 2.19.
Uma vez conhecida a impedância vista a partir da membrana timpânica, ZiT, pode facilmente
calcular-se as funções de transferência PT/PPW, PEC/PPW e PEX/PPW. Pode igualmente
determinar-se o circuito equivalente de Thévenin correspondente ao ouvido externo. O
equivalente de Thévenin referente à parte do modelo relativa à difracção do som consiste na
pressão e impedância equivalentes:
P PR sM
R sMd PWs s
s s
=++2
e ( ) ( )Z R sM R sMd s s r r// //= + . (2.64)
Considerando o produto das duas matrizes de parâmetros de transmissão, temos:
P
U
A B
C D
P
UEX
EX
T
T
=
⋅
, (2.65)
pelo que a pressão e impedância equivalentes de Thévenin do modelo do ouvido externo
são dadas por:
PP
A CZeqd
d
=+
e ZB DZ
A CZeqd
d
=++
. (2.66)
2.5.2 Ouvido médio
O modelo de Rosowski do ouvido médio é representado na figura 2.20. Uma descrição
pormenorizada das suas partes constituintes pode ser encontrada em [Rosowski, 96]. Este
modelo é utilizado neste trabalho com ligeiras alterações. Por exemplo, a impedância de
entrada da cóclea, ZC, é alterada para o paralelo de uma resistência, RC com a série de uma
resistência RC0 com uma indutância LC, em vez da ligação série de RC com LC representada
na figura 2.20. Esta alteração está em conformidade tanto com o valor teórico obtido
anteriormente como com valores experimentais.

42 Capítulo 2 - Modelos Cocleares
PC
Zm(a)
Pi = HmPPW
Zin(a)
US
a) b)
Figura 2.20 - a) Modelo do ouvido médio (in [Rosowski, 96]). PT representa a pressão timpânica, PS
a pressão efectiva que actua no estribo e PC a diferença de pressões entre as escalas timpânica evestibular. ATM é a área da membrana timpânica, AFP a área da base do estribo e (lM/lI) é o ganho dacadeia ossicular que resulta da rotação dos ossículos. Os valores usados dos parâmetros são osseguintes: CTC=4x10-12 m5N-1; CMC=3.9x10-11 m5N-1; RA=6x106 Pa s m-3; LA=100 Kg m-4; LT1=750Kg m-4; CT = 3x10-12 m5N-1; RT=3x107 Pa s m-3; LT=6.6x103 Kg m-4; CT2 =1.3x10-11 m5N-1;RT2=1.2x107 Pa s m-3; ATM=60x10-6 m-2; AFP=3.2x10-6 m-2; lM/lI =1.3; CTS=5x10-3 mN-1; RTS=4.3x10-2
Nsm-1; CMI=10-3 mN-1; RMI=7.2x10-3 Nsm-1; LMI=7.9x10-6 Kg; RJ=3.6 Nsm-1; CJ=4.9x10-4 mN-1;LS=3x10-6 Kg; CAL=5x10-14 m5N-1; RAL=0. Como impedância da cóclea considera-se o paralelo deuma resistência RC=1.55x1011 Pa s m-3; com a série de uma indutância, LC=8x107 Kg m-4 e umaresistência RC0=2.2x1010 Pa s m-3. b) Equivalente de Thévenin do ouvido externo e médio referido àentrada da cóclea, com pressão equivalente de entrada Pi e impedância acústica equivalente Zm
(a).
O valor de CAL, o inverso da rigidez atribuída às membranas das janelas oval, é também
alterado de forma a que a impedância acústica vista a partir do estribo, ZAL+ZC, apresenta
um mínimo a algumas dezenas de Hz, de acordo com medições experimentais [Lynch et al,
82]. A pressão PC corresponde assim à diferença de pressões entre as escalas na base da
cóclea. Os valores tomados para o modelo são indicados na legenda da figura 2.20 e
correspondem, na sua quase totalidade, aos valores originalmente propostos.
2.5.3 Resultados relativos ao ouvido externo e médio
A impedância vista a partir do tímpano pode agora ser calculada, bem como o equivalente
de Thévenin referente ao ouvido externo e médio, tal como se representa na figura 2.20b).
Os resultados mais importantes deste modelo indicam-se na figura 2.21. O ganho de pressão
à entrada da cóclea, PC/PPW, apresenta uma variação com a frequência que segue
aproximadamente o inverso da curva de limiar de audição. O ganho PC/PT está
razoavelmente de acordo com os dados obtidos em [Puria et al, 93]. O ganho de pressão
relativo ao modelo das cavidades do ouvido externo, PT/PPW, é genericamente correcto
[Chan & Geisler, 90] e coincide também razoavelmente com as medidas efectuadas em
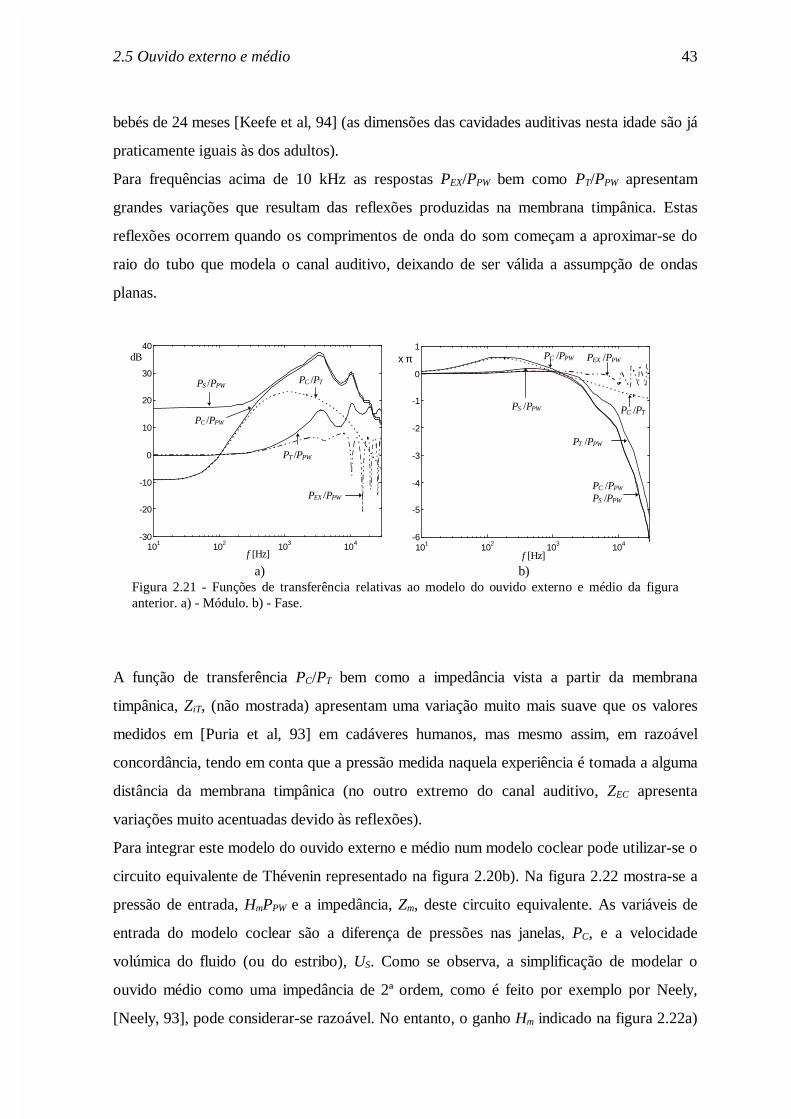
2.5 Ouvido externo e médio 43
bebés de 24 meses [Keefe et al, 94] (as dimensões das cavidades auditivas nesta idade são já
praticamente iguais às dos adultos).
Para frequências acima de 10 kHz as respostas PEX/PPW bem como PT/PPW apresentam
grandes variações que resultam das reflexões produzidas na membrana timpânica. Estas
reflexões ocorrem quando os comprimentos de onda do som começam a aproximar-se do
raio do tubo que modela o canal auditivo, deixando de ser válida a assumpção de ondas
planas.
101 102 103 104
-30
-20
-10
0
10
20
30
40
f [Hz]
dB
PC /PPW
PS /PPW
PT /PPW
PC /PT
PEX /PPW
101 102 103 104
-6
-5
-4
-3
-2
-1
0
1
f [Hz]
x π PC /PPW
PS /PPW
PT /PPW
PC /PT
PEX /PPW
PC /PPW
PS /PPW
a) b)Figura 2.21 - Funções de transferência relativas ao modelo do ouvido externo e médio da figuraanterior. a) - Módulo. b) - Fase.
A função de transferência PC/PT bem como a impedância vista a partir da membrana
timpânica, ZiT, (não mostrada) apresentam uma variação muito mais suave que os valores
medidos em [Puria et al, 93] em cadáveres humanos, mas mesmo assim, em razoável
concordância, tendo em conta que a pressão medida naquela experiência é tomada a alguma
distância da membrana timpânica (no outro extremo do canal auditivo, ZEC apresenta
variações muito acentuadas devido às reflexões).
Para integrar este modelo do ouvido externo e médio num modelo coclear pode utilizar-se o
circuito equivalente de Thévenin representado na figura 2.20b). Na figura 2.22 mostra-se a
pressão de entrada, HmPPW e a impedância, Zm, deste circuito equivalente. As variáveis de
entrada do modelo coclear são a diferença de pressões nas janelas, PC, e a velocidade
volúmica do fluido (ou do estribo), US. Como se observa, a simplificação de modelar o
ouvido médio como uma impedância de 2ª ordem, como é feito por exemplo por Neely,
[Neely, 93], pode considerar-se razoável. No entanto, o ganho Hm indicado na figura 2.22a)
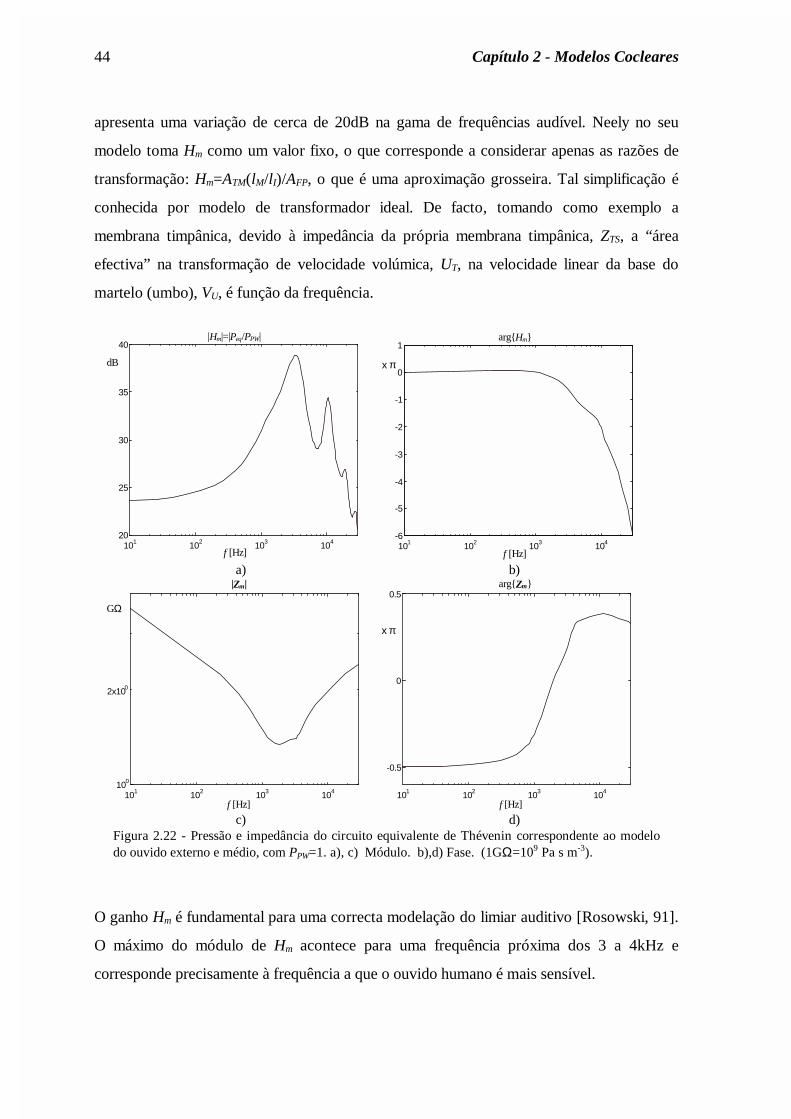
44 Capítulo 2 - Modelos Cocleares
apresenta uma variação de cerca de 20dB na gama de frequências audível. Neely no seu
modelo toma Hm como um valor fixo, o que corresponde a considerar apenas as razões de
transformação: Hm=ATM(lM/lI)/AFP, o que é uma aproximação grosseira. Tal simplificação é
conhecida por modelo de transformador ideal. De facto, tomando como exemplo a
membrana timpânica, devido à impedância da própria membrana timpânica, ZTS, a “área
efectiva” na transformação de velocidade volúmica, UT, na velocidade linear da base do
martelo (umbo), VU, é função da frequência.
101 102 103 104
20
25
30
35
40
f [Hz]
dB
|Hm|=|Peq/PPW|
101 102 103 104-6
-5
-4
-3
-2
-1
0
1
f [Hz]
x π
argHm
a) b)
101 102 103 104
100
f [Hz]
GΩ
|Zm|
2x100
101 102 103 104
-0.5
0
0.5
x π
argZm
f [Hz]
c) d)Figura 2.22 - Pressão e impedância do circuito equivalente de Thévenin correspondente ao modelodo ouvido externo e médio, com PPW=1. a), c) Módulo. b),d) Fase. (1GΩ=109 Pa s m-3).
O ganho Hm é fundamental para uma correcta modelação do limiar auditivo [Rosowski, 91].
O máximo do módulo de Hm acontece para uma frequência próxima dos 3 a 4kHz e
corresponde precisamente à frequência a que o ouvido humano é mais sensível.

2.6 Modelos Micromecânicos 45
Em conclusão, o modelo descrito do ouvido externo e médio pode considerar-se
suficientemente rigoroso simulando com algum detalhe dados resultantes de experiências
em humanos e animais. Trata-se de um modelo linear não entrando em conta com
fenómenos como histerese, reflexo acústico [Giguère & Woodland, 94b], etc., mas estes
fenómenos não lineares são pouco significativos a baixos níveis de excitação em ouvidos
normais.
Nas simulações do modelo coclear apresentadas no capítulo 3 este modelo é utilizado
através da integração do ganho Hm e impedância Zm no modelo de linha de transmissão da
figura 2.5.
2.6 Modelos Micromecânicos
Na década de 70 começaram a emergir dados que mostravam uma marcada não linearidade
das respostas cocleares em função do nível de excitação [Rhode, 71]. Um exemplo de não
linearidade consiste no decrescimento do pico das respostas cocleares com o nível de
excitação [Rhode, 71], [Sellick et al, 82]. Entretanto, outros fenómenos não lineares eram já
conhecidos utilizando como estímulo um par de tons sinusoidais. Um destes fenómenos é
conhecido como “supressão de dois tons”10 e consiste na redução da taxa de disparos de
uma fibra do nervo coclear devido à presença de um segundo tom [Sachs & Kiang, 68],
[Sachs & Abbas, 74, 76], [Delgutte, 90], [Ruggero et al, 92], [Nuttal & Dolan, 93]. Outro
fenómeno não linear consiste na geração de tons adicionais (aos dois tons de estímulo) de
combinação, fenómeno este também conhecido como produtos de distorção de inter-
modulação ou simplesmente como produtos de distorção. Consiste na produção de tons
relacionados com os tons do estímulo de frequências f1 e f2 na forma mf1-nf2 onde m e n são
números inteiros, sendo a distorção mais importante a chamada distorção cúbica, 2f2-f1, [de
Boer, 84], [Furst & Goldstein, 82], [Ruggero, 92], [Allen & Fahey, 93]. O estudo destes
tons de combinação, primeiro por estudos psico-físicos [Goldstein, 67] e depois por
experiências fisiológicas no nervo auditivo e cóclea [Goldstein & Kiang, 68], [Robles et al,
91], teve um papel determinante na teoria coclear moderna.
10 “Two-Tone Suppression” na designação original.

46 Capítulo 2 - Modelos Cocleares
Surgiram então modelos cocleares que tentavam descrever este comportamento não linear.
Os modelos eram baseados na formulação 1D mas necessariamente simulados no domínio
temporal. Na maior parte dos casos o factor de atrito ou resistência da impedância da
partição variava proporcionalmente com a velocidade da partição a fim de reduzir a
sensibilidade das respostas. Modelos deste tipo [Hall, 80], [Deng & Geisler, 87], [Zwicker,
86], [Fust & Goldstein, 82], [Strube, 85], etc., permitiram descrever, de uma forma mais ou
menos completa, o comportamento não linear das respostas cocleares e neuronais então
conhecidas. Contudo, o andamento das respostas continuava a não corresponder aos dados
mecânicos conhecidos tal como foi explicado através do modelo clássico abordado nas
secções anteriores.
Nesta altura os métodos de análise não estavam tão desenvolvidos como hoje e existiam
discrepâncias significativas entre dados mecânicos e dados neuronais. Uma explicação
plausível para esta discrepância fundamentava-se na suposição que a vibração medida por
todos os investigadores nessa altura não corresponderia à vibração que estimulava as células
sensoriais. Muito do esforço teórico que se seguiu tentou determinar se a vibração
micromecânica do órgão de Corti poderia explicar estas discrepâncias o que levou ao
surgimento da teoria do “segundo filtro”.
Membrana Tectória
Membrana Basilar
Células Ciliadas Externas
Célula Ciliada Interna
Fibras Nervosas
MB
MT
MB
KC
KT
z
y
Figura 2.23 - a) - Esquema do orgão de Corti. b) - Modelo mecânico simplificado. A membranabasilar move-se no sentido vertical e a membrana tectória desliza numa direcção radial reforçando aoscilação dos cílios das células ciliadas. Existem assim dois modos de vibração ou graus de liberdade.
A ideia base nesta abordagem é que uma segunda ressonância estaria envolvida na
impedância da partição e que teria a ver com a membrana tectória e com os cílios das
células ciliadas. Na figura 2.23 apresenta-se um esquema do órgão de Corti que permite
visualizar o princípio básio de funcionamento. Quando a membrana basilar se move no

2.6 Modelos Micromecânicos 47
sentido vertical, a membrana tectória pode rodar em torno da sua conexão óssea mas
também pode deslizar livremente numa direcção radial (direcção y na figura 2.23b). Existem
assim dois graus de liberdade na oscilação das estruturas do órgão de Corti. O movimento
radial da membrana tectória não interfere directamente com o movimento do fluido coclear,
mas permite uma oscilação dos cílios das CCEs e CCIs muito superior à que seria
conseguida apenas pelo movimento vertical da membrana basilar.
Um trabalho notável de modelação do órgão de Corti foi feita por Allen, [Allen, 80], que,
dada a sua importância, será abordado na secção seguinte.
Desde cedo que Dallos e seus colaboradores [Lyon & Mead, 88], [Dallos, 96], apontavam
para a necessidade de considerar as CCEs no funcionamento normal da cóclea, baseada na
redução muito significativa de sensibilidade das respostas neuronais quando as CCEs são
destruídas por antibióticos ototóxicos. Não era muito claro na altura como é que isto
poderia acontecer pois se sabia que as CCEs recebem apenas 5% dos neurónios aferentes e
estava amplamente estabelecido que a vibração coclear era passiva, linear e robusta
[Patuzzi, 96]. Porém, no início da década de 80 foi progressivamente tornando-se claro que
a cóclea teria de ser activa sendo este comportamento devido às CCEs [de Boer, 83, 83b,
91], [Davis, 83], [Kim, 84], [Zweig, 91], [Brass & Kemp, 93]. A descoberta da mobilidade
destas células (capacidade de expansão e contracção através de estimulação eléctrica até
cerca de 30kHz, [Brownell et al, 85], [Holley, 96]) permitiu consolidar esta hipótese.
Os modelos cocleares seguiram obviamente por esta via. De entre os modelos activos
recentemente propostos talvez os mais importantes sejam os modelos de Neely-Kim [Neely
& Kim, 86], de Geisler [Geisler, 91, 93] e de Neely [Neely, 93] e por isso mesmo serão
descritos com algum pormenor seguidamente. Muitos outros modelos têm vindo a ser
propostos nos últimos anos, [Neely & Stover, 93], [Geisler & Sang 95], [Mammano &
Nobili, 93], [Hubbard, 93], [de Boer, 93], etc., embora sejam aqui referidos apenas de
passagem.
2.6.1 Modelo de Allen
Na figura 2.24a) mostra-se o modelo mecânico proposto por Allen [Allen, 80]. Neste
modelo admite-se que as membranas basilar e tectória podem rodar segundo eixos situado

48 Capítulo 2 - Modelos Cocleares
junto à parede coclear. A membrana tectória pode deslocar-se radialmente constituindo um
2º grau de liberdade de movimento.
KB
MB
MT
RC KKT
MB
MT
LR
M RB KB
P
VBM
MT
RT
KTKC
RC
1/g
VTVC
a) b)Figura 2.24 - a) Esquema mecânico da partição coclear. MB: membrana basilar; MT: membranatectória. LR: lâmina reticular [Allen, 80]. b) Esquema eléctrico equivalente.
Allen considerou que no deslocamento da membrana basilar influíam tanto a massa da
membrana basilar como da membrana tectória, e não considerou a existência de atrito na
membrana basilar (embora esteja representado no esquema da figura 2.24b)). Deixando de
parte alguns pormenores, identificam-se neste esquema 3 impedâncias: impedância da
membrana basilar, da membrana tectória e ainda uma impedância de acoplamento entre as
duas membranas. Não é evidente neste esquema o ganho de deslocamento radial face ao
deslocamento vertical da membrana basilar, g. O esquema eléctrico correspondente
representa-se na figura 2.24b), onde P representa a diferença de pressões através da
membrana basilar; VBM e VT representam as velocidades lineares das membranas basilar e
tectória, respectivamente, e VC a velocidade de deslizamento entre as duas membranas, a
que se associa a velocidade de deformação dos cílios das células ciliadas.
Contrariamente ao modelo clássico, onde se considerava que o órgão de Corti se movia
solidariamente com a membrana basilar e, portanto, se considerava apenas uma impedância,
agora a impedância da partição apresenta um segundo termo devido ao 2º grau de
liberdade. A impedância da partição é agora:
ZP
VZ g Z //Zp
BMBM C T= = + =2 ( )
= + + ++ + +
+ + + +sM R
K
sg
R K s s M sR K
s M s R R K KBB C C T T T
T C T T C
22
2
( )( )
( ) ( ). (2.67)

2.6 Modelos Micromecânicos 49
Aliás, este esquema em que a impedância da partição é a soma da impedância da membrana
basilar com uma outra impedância devida ao órgão de Corti, é seguido em todos os
modelos micromecânicos lineares recentes. Neste caso, a impedância descrita pela equação
anterior é uma impedância de 4ª ordem, definida com 4 zeros e 3 pólos, um dos quais na
origem.
Allen admitiu que poderia ser a velocidade ciliar, VC, a grandeza responsável pelos disparos
das CCIs (células que têm ligações aferentes: transmitem informação da cóclea para o
núcleo coclear). Desta forma, a relação entre a velocidade VC e a velocidade da membrana
basilar explica a diferença (então) observada entre os dados mecânicos referentes à partição
coclear e os dados neuronais, isto é, constituía o procurado “segundo filtro”. Este segundo
filtro corresponde à seguinte função de transferência de 2ª ordem:
V
VH g
Z
Z Zg
s sR
M
K
M
s sR R
M
K K
M
C
BMT
T
T C
T
T
T
T
T C
T
T C
T
= =+
=+ +
++
++
2
2
. (2.68)
Esta função pode ser estimada através dos parâmetros físicos em jogo mas também através
dos dados neuronais uma vez que HT se pode colocar na forma seguinte:
H x s gs d
s dTz z z
p p p
( , ) =+ ++ +
2 2
2 2
ω ωω ω
. (2.69)
A frequência ωz deveria estar aproximadamente uma oitava abaixo de ωCF de forma a
explicar a existência de um zero espectral observado nas curvas de sintonização neuronal.
ωp deverá situar-se ligeiramente acima de ωCF, a frequência característica tonotópica [Allen,
80]. Esta característica era já explorada num modelo fenomenológico11 anterior [Allen, 77].
Na figura 2.25 mostra-se a impedância da partição segundo este modelo bem como a
velocidade VC obtida com o modelo 1D. Como se pode observar, VC apresenta uma
pequena depressão a cerca de metade da frequência característica que resulta do zero
espectral em HT.
11 O termo fenomenológico aplica-se a modelos que simulam um dado fenómeno conhecido sem no entantodescrever os processos físicos ou fisiológicos envolvidos.
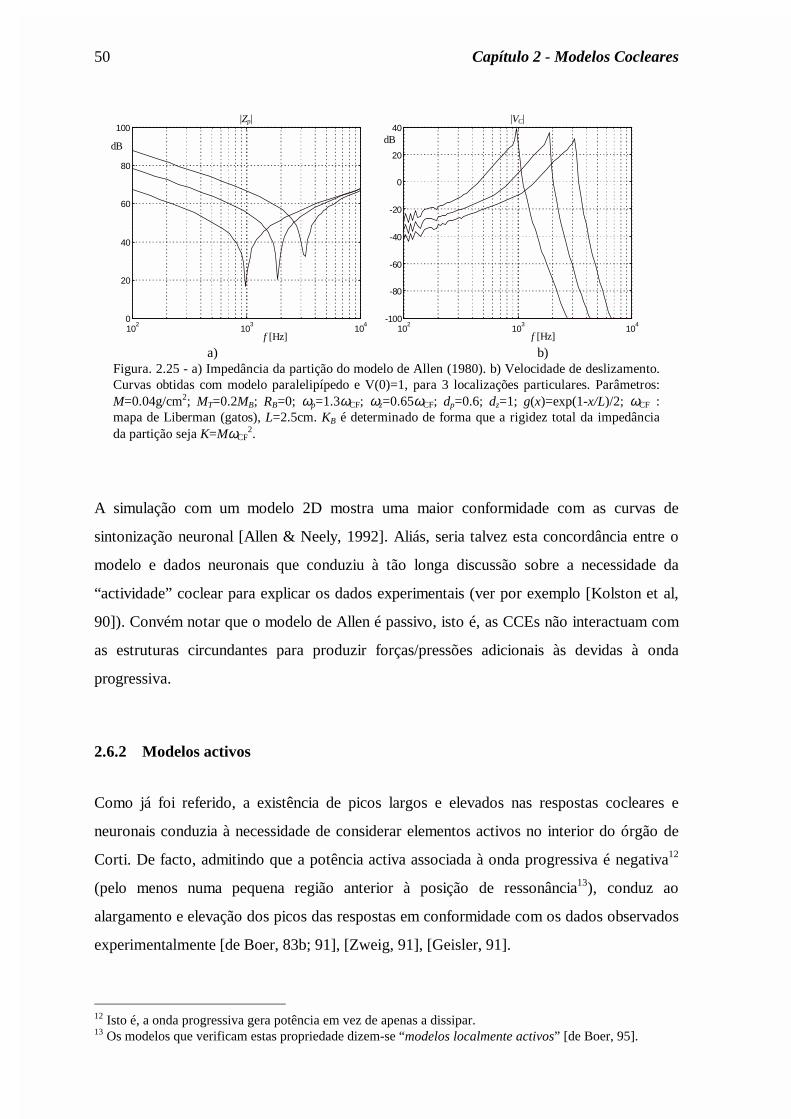
50 Capítulo 2 - Modelos Cocleares
10
210
310
40
20
40
60
80
100
f [Hz]
dB
|Zp|
10
210
310
4-100
-80
-60
-40
-20
0
20
40
f [Hz]
dB
|VC|
a) b)Figura. 2.25 - a) Impedância da partição do modelo de Allen (1980). b) Velocidade de deslizamento.Curvas obtidas com modelo paralelipípedo e V(0)=1, para 3 localizações particulares. Parâmetros:M=0.04g/cm2; MT=0.2MB; RB=0; ωp=1.3ωCF; ωz=0.65ωCF; dp=0.6; dz=1; g(x)=exp(1-x/L)/2; ωCF :mapa de Liberman (gatos), L=2.5cm. KB é determinado de forma que a rigidez total da impedânciada partição seja K=MωCF
2.
A simulação com um modelo 2D mostra uma maior conformidade com as curvas de
sintonização neuronal [Allen & Neely, 1992]. Aliás, seria talvez esta concordância entre o
modelo e dados neuronais que conduziu à tão longa discussão sobre a necessidade da
“actividade” coclear para explicar os dados experimentais (ver por exemplo [Kolston et al,
90]). Convém notar que o modelo de Allen é passivo, isto é, as CCEs não interactuam com
as estruturas circundantes para produzir forças/pressões adicionais às devidas à onda
progressiva.
2.6.2 Modelos activos
Como já foi referido, a existência de picos largos e elevados nas respostas cocleares e
neuronais conduzia à necessidade de considerar elementos activos no interior do órgão de
Corti. De facto, admitindo que a potência activa associada à onda progressiva é negativa12
(pelo menos numa pequena região anterior à posição de ressonância13), conduz ao
alargamento e elevação dos picos das respostas em conformidade com os dados observados
experimentalmente [de Boer, 83b; 91], [Zweig, 91], [Geisler, 91].
12 Isto é, a onda progressiva gera potência em vez de apenas a dissipar.13 Os modelos que verificam estas propriedade dizem-se “modelos localmente activos” [de Boer, 95].

2.6 Modelos Micromecânicos 51
Os modelos cocleares activos funcionam de acordo com esta propriedade. Um dos
primeiros modelos activos a ser proposto foi o modelo de Neely-Kim. Outros mais
elaborados se seguiram, nomeadamente os modelos de Geisler, de Kanis-deBoer e de
Neely. Todos estes modelos vão ser abordados seguidamente.
2.6.21. Modelo de Neely-Kim
O modelo de Neely-Kim [Neely & Kim, 86], coincide nos pressupostos mecânicos com o
modelo de Allen, antes focado, mas as CCEs ao alongarem-se e a contraírem-se conduzem
ao aparecimento de uma pressão que vai reforçar a diferença de pressões através da
partição e com isso fortalecer a onda progressiva. O esquema equivalente eléctrico deste
modelo está representado na figura 2.26. A pressão activa, Pa, depende da velocidade ciliar
da seguinte forma:
P gZ Va NK C= −γ , (2.70)
onde γ é um parâmetro de controlo do ganho de “actividade” do modelo: com γ=0 o
modelo é passivo e com γ=1 teremos a máxima actividade com o sistema ainda estável. A
impedância de transferência ZNK controla a distribuição da actividade com a posição coclear
e frequência e é definida com um termo resistivo e um termo de rigidez.
MB RB KB
P
VBM
MT
RT
KTKC
RC
1/g
VTVC
Pa
ZBM
g2(ZT//ZC)P-Pa
VBM
+−
+
a) b)Figura 2.26 - a) Esquema equivalente do modelo de Neely-Kim. b) Circuito reduzido ao primário.
Relativamente ao modelo de Allen (ver (2.68)), neste modelo a impedância da partição
apresenta mais um termo devido à pressão Pa:
Z Z g Z Z gZ Hp BM C T NK T= + −2 ( / / ) γ , (2.71a)
ou
[ ]Z Z gH Z Zp BM T C NK= + −γ . (2.71b)
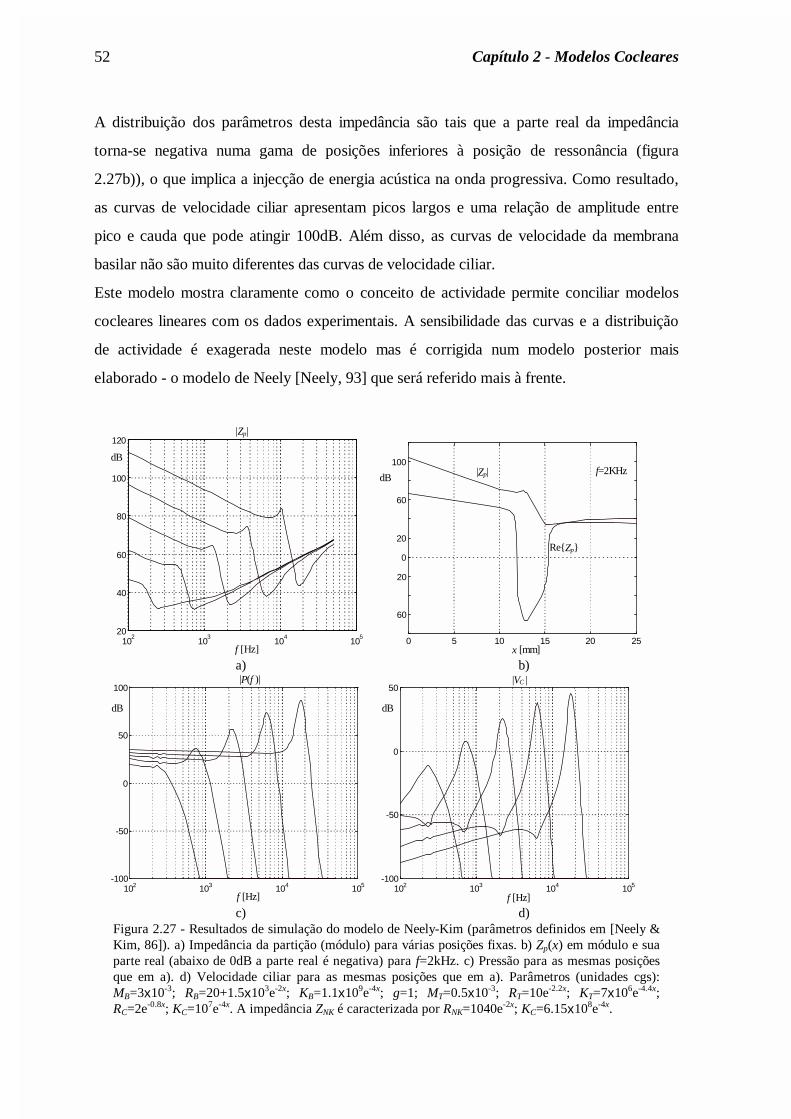
52 Capítulo 2 - Modelos Cocleares
A distribuição dos parâmetros desta impedância são tais que a parte real da impedância
torna-se negativa numa gama de posições inferiores à posição de ressonância (figura
2.27b)), o que implica a injecção de energia acústica na onda progressiva. Como resultado,
as curvas de velocidade ciliar apresentam picos largos e uma relação de amplitude entre
pico e cauda que pode atingir 100dB. Além disso, as curvas de velocidade da membrana
basilar não são muito diferentes das curvas de velocidade ciliar.
Este modelo mostra claramente como o conceito de actividade permite conciliar modelos
cocleares lineares com os dados experimentais. A sensibilidade das curvas e a distribuição
de actividade é exagerada neste modelo mas é corrigida num modelo posterior mais
elaborado - o modelo de Neely [Neely, 93] que será referido mais à frente.
10
210
310
410
520
40
60
80
100
120
f [Hz]
dB
|Zp|
0 5 10 15 20 25
60
20
0
20
60
100
x [mm]
dB|Zp|
ReZp
f=2KHz
a) b)
102 103 104 105
-100
-50
0
50
100
f [Hz]
dB
|P(f )|
102 103 104 105
-100
-50
0
50
f [Hz]
dB
|VC |
c) d)Figura 2.27 - Resultados de simulação do modelo de Neely-Kim (parâmetros definidos em [Neely &Kim, 86]). a) Impedância da partição (módulo) para várias posições fixas. b) Zp(x) em módulo e suaparte real (abaixo de 0dB a parte real é negativa) para f=2kHz. c) Pressão para as mesmas posiçõesque em a). d) Velocidade ciliar para as mesmas posições que em a). Parâmetros (unidades cgs):MB=3x10-3; RB=20+1.5x103e-2x; KB=1.1x109e-4x; g=1; MT=0.5x10-3; RT=10e-2.2x; KT=7x106e-4.4x;RC=2e-0.8x; KC=107e-4x. A impedância ZNK é caracterizada por RNK=1040e-2x; KC=6.15x108e-4x.

2.6 Modelos Micromecânicos 53
2.6.22. Modelo de Kanis-deBoer
Trata-se de um modelo coclear adaptado directamente do modelo clássico abordado
anteriormente, mas onde a impedância da partição é activa, embora seja definida de uma
forma fenomenológica. Neste modelo [Kanis & de Boer, 93; 94], admite-se, tal como no
modelo de Neely-Kim, que as CCEs geram uma pressão que é adicionada à diferença de
pressões através da partição. A impedância da partição tem assim mais um termo, ZOC,
associado ao órgão de Corti. O efeito de ZOC na impedância da partição consiste em tornar
a parte real negativa abaixo da frequência de ressonância, de acordo com análise teórica
sobre a amplitude das respostas cocleares [de Boer, 83]. A definição das impedâncias é a
seguinte:
Z Z Zp BM OC= − (2.72)
Zs s
s sOCSC
=
++ +
γω ω
δ ω σ ω 0 0
20
2020 06.
( ) . (2.73)
A impedância relativa ao órgão de Corti é assim postulada e não deriva portanto de uma
relação física explícita. Nesta última expressão ω0 corresponde à frequência de ressonância
a variar exponencialmente com x (expressão (2.40)). O parâmetro γ controla a actividade do
modelo (para γ >1 o modelo torna-se instável). Os parâmetros δSC e σ são constantes com x
(δSC=0,14; σ=0,7) e definem um par de pólos complexos nesta impedância de 2ª ordem.
Com esta definição, a impedância da partição apresenta a mesma simetria de escala referida
na secção 2.3.2. A impedância da membrana basilar é defina com os parâmetros do modelo
clássico indicados na figura 2.7, mas com um factor de amortecimento δ muito superior:
δ=0.4. Assim, com γ=1 a impedância da partição pode ser colocada na seguinte forma:
( ) ( )( )Z u x
M x
uu d
u ud
u udp Za Za Za
az az az
ap ap ap
( , )( )
= + ++ +
+ +
ω0 2 2
2 2
2 2Ω Ω
Ω Ω
Ω Ω, (2.74)
onde u é a frequência normalizada, u=s/ω0. As frequências de ressonância normalizadas e os
factores de amortecimento são as seguintes: ΩZa=0,9017; dZa=0.029; Ωaz=0,7763;
daz=0.507, dapΩap=δSC, Ωap=ΩZaΩaz=σ=0,7. Note-se o baixo factor de amortecimento dZa
(factor de qualidade QZa=34) em relação a δ que faz com que o 2º factor na expressão
anterior domine na zona de ressonância. Pode verificar-se que devido a esta propriedade de
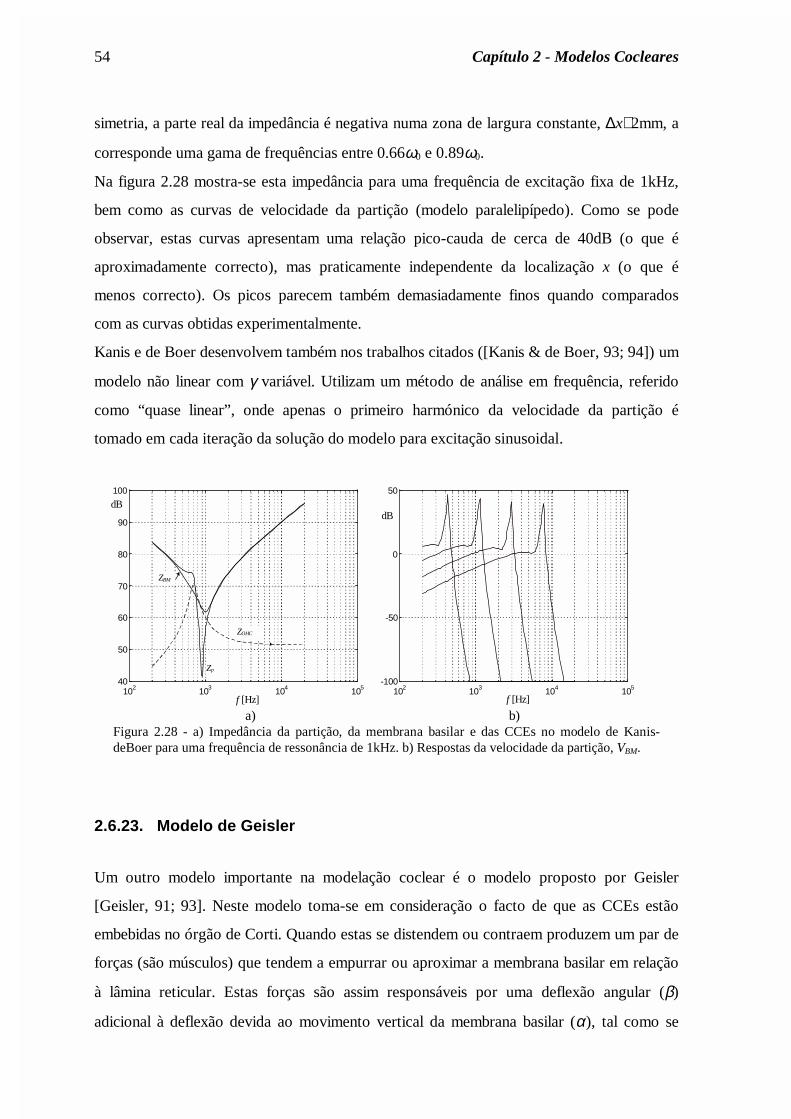
54 Capítulo 2 - Modelos Cocleares
simetria, a parte real da impedância é negativa numa zona de largura constante, ∆x≅ 2mm, a
corresponde uma gama de frequências entre 0.66ω0 e 0.89ω0.
Na figura 2.28 mostra-se esta impedância para uma frequência de excitação fixa de 1kHz,
bem como as curvas de velocidade da partição (modelo paralelipípedo). Como se pode
observar, estas curvas apresentam uma relação pico-cauda de cerca de 40dB (o que é
aproximadamente correcto), mas praticamente independente da localização x (o que é
menos correcto). Os picos parecem também demasiadamente finos quando comparados
com as curvas obtidas experimentalmente.
Kanis e de Boer desenvolvem também nos trabalhos citados ([Kanis & de Boer, 93; 94]) um
modelo não linear com γ variável. Utilizam um método de análise em frequência, referido
como “quase linear”, onde apenas o primeiro harmónico da velocidade da partição é
tomado em cada iteração da solução do modelo para excitação sinusoidal.
102 103 104 105
40
50
60
70
80
90
100
f [Hz]
dB
ZOHC
Zp
ZBM
102 103 104 105
-100
-50
0
50
f [Hz]
dB
a) b)Figura 2.28 - a) Impedância da partição, da membrana basilar e das CCEs no modelo de Kanis-deBoer para uma frequência de ressonância de 1kHz. b) Respostas da velocidade da partição, VBM.
2.6.23. Modelo de Geisler
Um outro modelo importante na modelação coclear é o modelo proposto por Geisler
[Geisler, 91; 93]. Neste modelo toma-se em consideração o facto de que as CCEs estão
embebidas no órgão de Corti. Quando estas se distendem ou contraem produzem um par de
forças (são músculos) que tendem a empurrar ou aproximar a membrana basilar em relação
à lâmina reticular. Estas forças são assim responsáveis por uma deflexão angular (β)
adicional à deflexão devida ao movimento vertical da membrana basilar (α), tal como se

2.6 Modelos Micromecânicos 55
representa na figura 2.29. A membrana tectória segue o deslocamento da lâmina reticular e
os cílios das CCEs sofrem uma deflexão aproximadamente proporcional a α+β.
MT
pilaresx
LR z
MB
y
CCE
α
β
Figura 2.29 - Esquema geométrico das forças e deslocamentos envolvidos na descrição do modelo deGeisler. MB- membrana basilar; MT - membrana tectória, LR - lâmina reticular, CCE- célula ciliadaexterna. Geisler considera os deslocamentos ma membrana basilar (x),da lâmina reticular (z) e adeflexão dos cílios (y).
Geisler toma em conta basicamente o modelo geométrico representado na figura anterior e
considera o deslocamento produzido pela membrana basilar, x, pela lâmina reticular, z, e a
deflexão dos cílios das CCEs, y. A estes deslocamentos podem associar-se as respectivas
velocidades, VBM, Vr e Vc. À velocidade Vc está associada a impedância Zc que consiste no
paralelo da impedância da membrana tectória e a impedância de acoplamento dos cílios.
Existe ainda uma impedância, Zr, que representa a rigidez axial das CCEs e das células de
suporte. Uma análise detalhada das equações do modelo permite chegar ao modelo
equivalente representado na figura 2.30 14. Neste circuito, PCCE representa uma pressão
equivalente resultante da força provocada pelas CCEs e que se considera proporcional ao
deslocamento ciliar. Esta relação pode traduzir-se da seguinte forma:
P Z VCCE G c= γ . (2.75)
O parâmetro γ tem o mesmo significado que anteriormente. ZG é a impedância de
transferência que traduz a distribuição posição-frequência das forças associadas às CCEs. A
impedância da partição toma então a seguinte forma (eq. (4) em [Geisler, 93]):
14 Para chegar a este circuito equivalente é necessário fazer a correspondência entre os deslocamentos x, y ez e as velocidades VBM, Vc e Vr tendo em conta a geometria do modelo. Fazendo uma análise cuidadosa dasequações do modelo em [Geisler, 93], pode mostrar-se que esta correspondência é a seguinte:
VBM=sx; Vc=gsy/AE; Vr = −gszB/A, PCCE=VcFfb/s = γZGVc ,onde s=jω e g, A, B e E são constantes relacionadas com a geometria do modelo. A pressão PCCE é portantoproporcional ao deslocamento ciliar y, onde Ffb (ou ZG) traduz a distribuição posição-frequência das forçascontrácteis das CCEs. As constantes são determinadas a partir das seguintes relações: A/B=1.71;C=0.57cotan(32o); D = C+cos(18o); g2=DA/B. A constante E fica determinada através da definição de Ffb ouZG, definida mais adiante.

56 Capítulo 2 - Modelos Cocleares
Z Z gZ Z
Z Z ZZ Zp BM
c r
r c GBM OC= +
+ += +2
γ . (2.76)
P
VBM
Zr
1/g
Vc
Vr
PCCE
ZBM
+
+−
Zc
Figura 2.30 - Esquema equivalente do modelo de Geisler.
A constante g é determinada pela geometria do modelo, da mesma forma que no modelo de
Allen, mas é aqui independente da posição coclear. As impedâncias Zc e Zr são
caracterizadas apenas pelo termo de rigidez: Zc=Kc/s, Zr=Kr/s. A impedância da membrana
basilar é composta por massa (constante), resistência e rigidez da seguinte forma,
( )Z s sM R K sM
ss d x s xBM b b b
b( ) ( ) ( )= + + = + +20 0
2ω ω , (2.77)
onde d=0,6 e onde ω0(x) é a frequência característica tonotópica. Também neste caso foi
usado o mapa de Liberman. Finalmente, Geisler propõe a seguinte forma para a impedância
ZG:
ZK
s
s
seG
b r
r
sTr=−+
−ω ω
ω ω0
0
. (2.78)
Esta forma é motivada pelo trabalho teórico de Zweig [Zweig, 91] cujo resultado indicia a
existência de um atraso (Tr) na resposta das células ciliadas, inversamente proporcional à
frequência característica tonotópica: Tr=0,4π/ω0. A rigidez atribuída às CCEs apresenta um
valor próximo da rigidez da membrana basilar e depende portanto do valor de γ tomado.
Geisler refere o valor de 1,29 como valor óptimo de γ, e não deve exceder 1,58 por
questões de estabilidade. A expressão em parênteses rectos, onde ωr=ω0/20 rad/s,
corresponde uma filtragem passa-tudo e portanto provoca apenas uma variação de fase.
Esta impedância pode ser vista como o produto da impedância Zb=Kb/s por uma função de
transferência HG , passa-tudo, associada a uma filtragem (apenas desvio de fase) operada
pelas CCEs. A impedância da partição pode colocar-se portanto na seguinte forma:

2.6 Modelos Micromecânicos 57
Z Z gZ Z
HZ
Z Z
p BMc r
Gb
c r
= ++
+
2
1
/ /
γ . (2.79)
Também neste modelo a impedância da partição apresenta a mesma propriedade de simetria
referida anteriormente pois todos os parâmetros na definição da impedância são referidos a
ω0. Verifica-se que para frequências acima de, cerca de duas vezes a frequência
característica, o valor de HG deixa de ser importante, pois a impedância da membrana
basilar passa a dominar. De acordo com dados experimentais recentes esta função de
transferência HG deveria apresentar uma característica passa-baixo. Geisler refere que
incorporando uma função passa-baixo no modelo resulta em picos das respostas com
amplitudes demasiado baixas. Contudo, pelo exposto, é possível fazer com que HG
apresente um comportamento passa-baixo acima de aproximadamente 2ω0, sem que isto
altere significativamente a impedância da partição. Pode, por exemplo, utilizar-se uma
aproximação racional da função HG, com módulo unitário (ou aproximadamente unitário)
até 2ω0(x) e com aproximadamente a mesma fase. Assim, para uma frequência normalizada
u=jω/ω0(x), a função HG pode aproximar-se facilmente por uma função racional na forma:
H uB u
A uuG ( )
( )
( ), |≅ < | 2 , (2.80)
onde B e A são polinómios em u. Desta forma, é possível associar a HG uma função passa-
baixo (ou passa-tudo) com frequência de corte de cerca de 2ω0 (u≈2). Resulta assim a
seguinte aproximação racional para a impedância Zp:
( )Z uM x
uu ud g
K //K A u
A uK
K KB u
pb r c
b
r c
(mod) ( )( ) ( ) ( )
( ) ( )= + + +
++
ω
γ
0 2 21
. (2.81)
Por exemplo, com A(u) de grau 4 e B(u) de grau 3, podem obter-se os coeficientes dos
polinómios por um processo de minimização quadrática resultando uma função HG passa
baixo, para a qual a impedância da partição é praticamente indistinguível da impedância
original. Tal pode ser observado na figura 2.31 a) e b). Nessa figura apresenta-se a
impedância da partição bem como as suas componentes, ZBM e ZOC. As aproximações
racionais Zp(rac) e correspondente ZOC
(rac) estão também representadas. De facto, a
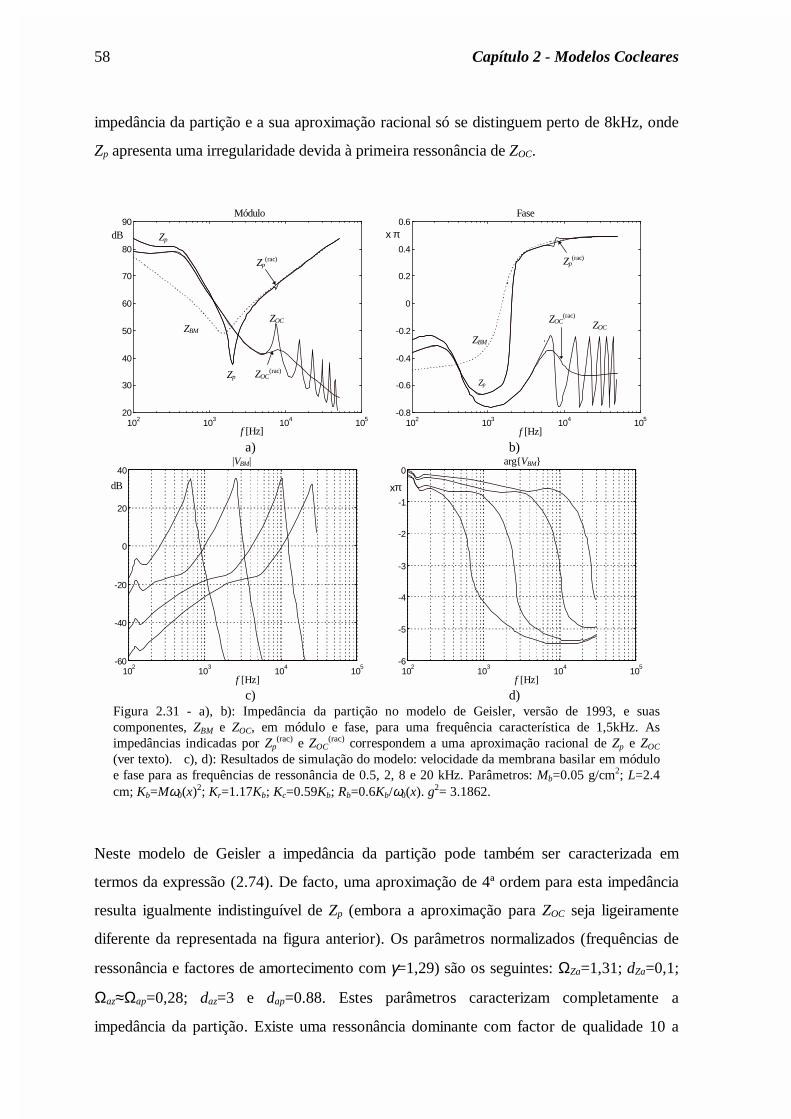
58 Capítulo 2 - Modelos Cocleares
impedância da partição e a sua aproximação racional só se distinguem perto de 8kHz, onde
Zp apresenta uma irregularidade devida à primeira ressonância de ZOC.
ZOC(rac)
ZOC
102 103 104 10520
30
40
50
60
70
80
90Módulo
f [Hz]
dB
ZBM
Zp
Zp(rac)
Zp
ZBM
Zp
ZOC
102 103 104 105-0.8
-0.6
-0.4
-0.2
0
0.2
0.4
0.6Fase
f [Hz]
x π
ZOC(rac)
Zp(rac)
a) b)
10
210
310
410
5-60
-40
-20
0
20
40|VBM|
f [Hz]
dB
10
210
310
410
5-6
-5
-4
-3
-2
-1
0argVBM
f [Hz]
xπ
c) d)Figura 2.31 - a), b): Impedância da partição no modelo de Geisler, versão de 1993, e suascomponentes, ZBM e ZOC, em módulo e fase, para uma frequência característica de 1,5kHz. Asimpedâncias indicadas por Zp
(rac) e ZOC(rac) correspondem a uma aproximação racional de Zp e ZOC
(ver texto). c), d): Resultados de simulação do modelo: velocidade da membrana basilar em móduloe fase para as frequências de ressonância de 0.5, 2, 8 e 20 kHz. Parâmetros: Mb=0.05 g/cm2; L=2.4cm; Kb=Mω0(x)2; Kr=1.17Kb; Kc=0.59Kb; Rb=0.6Kb/ω0(x). g2= 3.1862.
Neste modelo de Geisler a impedância da partição pode também ser caracterizada em
termos da expressão (2.74). De facto, uma aproximação de 4ª ordem para esta impedância
resulta igualmente indistinguível de Zp (embora a aproximação para ZOC seja ligeiramente
diferente da representada na figura anterior). Os parâmetros normalizados (frequências de
ressonância e factores de amortecimento com γ=1,29) são os seguintes: ΩZa=1,31; dZa=0,1;
Ωaz≈Ωap=0,28; daz=3 e dap=0.88. Estes parâmetros caracterizam completamente a
impedância da partição. Existe uma ressonância dominante com factor de qualidade 10 a

2.6 Modelos Micromecânicos 59
uma frequência cerca de 1,3ω0 e um par de pólos e de zeros praticamente à mesma
frequência, a cerca de um quarto de ω0, mas com o factor de qualidade do par de pólos
dominante. Na figura 2.31 c) e d) apresenta-se a velocidade da membrana basilar obtida
com o modelo paralelipípedo 1D. Este modelo mostra que não é necessário a existência de
uma segunda ressonância associada à membrana tectória para que os resultados mostrem
uma razoável concordância com dados fisiológicos. De facto, a relação entre as velocidades
Vc e VBM em função da frequência não mostra a existência de qualquer ressonância. Uma
deficiência do modelo é que o abaixamento da actividade (com γ menor) não conduz, como
deveria, a um abaixamento significativo da frequência dos picos das respostas, aliás como
também não acontece no modelo de Kanis-deBoer.
2.6.24. Modelo de Neely
Um dos modelos lineares mais completos propostos até hoje é o modelo de Neely [Neely,
93]. Parte do mesmo princípio que o modelo de Geisler, isto é, que o efeito das CCEs
consiste em separar a membrana basilar da lâmina reticular. Contudo, este efeito é tomado
de uma forma extrema: a separação entre a lâmina reticular e a membrana basilar é
independente da resistência que encontra. A velocidade (vertical) de contracção das CCEs,
Vc, considera-se proporcional à velocidade de deflexão lateral dos cílios, V0, e depende de
uma função de ganho, Hc, que representa a transdução operada pelas CCEs: Vc=γHcV0. Em
termos eléctricos, isto significa que em vez de uma fonte de tensão a fonte activa consiste
agora numa fonte de corrente. O circuito equivalente para este modelo é o que se apresenta
na figura 2.32 (figura 4 em [Neely, 93]).
P
VBM
Zt
1/g
V0
Vt
Vc =γHcV0
ZBM
+
Z0
Vr
Figura 2.32 - Esquema equivalente do modelo de Neely. Zt: impedância da membrana tectória; Zo:impedância de acoplamento entre a membrana basilar e a lâmina reticular. O transformador separaas vibrações tomadas verticalmente das vibrações laterais.

60 Capítulo 2 - Modelos Cocleares
A filtragem operada pelas CCEs tem duas componentes (ambas filtros passa baixo de 1ª
ordem). Uma que corresponde à transdução mecânico-eléctrica (correspondente às ligações
aferentes ou directas das fibras no nervo auditivo) e outra electromecânica (ligações
eferentes ou reversas). A função de transferência Hc tem portanto a seguinte tradução:
( )( )Hg
s sc
fr f r
f r
=+ +
ω ω
ω ω , (2.82)
onde os parâmetros gfr, ωf e ωr , dependem da posição coclear. Estes parâmetros são
escolhidos de forma a que os resultados de simulação do modelo sejam o mais próximo
possível das curvas obtidas de forma experimental.
Neste modelo faz-se também uma descrição do funcionamento das CCIs. Assume-se que os
cílios das CCIs não estão em contacto com a membrana tectória e que portanto são
deflectidos pelo fluido por arrastamento lateral. A deflexão dos cílios toma-se proporcional
ao deslocamento do fluido a altas frequências e à velocidade do fluido para baixas
frequências. A frequência de transição é ωi (2πx1390 rad.). Assume-se também que o
deslizamento entre a membrana tectória e lâmina reticular (Vr =g(VBM−Vc)), perto das CCIs,
é menos afectado (factor α) pela contracção das CCE’s (Vc) que quando perto das CCEs.
Assim, a velocidade de deflexão dos cílios das CCIs é calculado da seguinte forma15:
( )( )Vs
sg V V V H Vi
iBM c t i BM=
+− − − = ⋅
ωα( )1 . (2.83)
A relação entre a velocidade (ou deslocamento) dos cílios das CCEs com a velocidade
(deslocamento) da membrana basilar é:
HV
V
gH
gH HH
Z
Z ZBM
t
c tt
t
t0
0 0
00
01= =
+=
+γ , , (2.84)
enquanto para as CCIs a relação é:
Hs
sH gHi
ic=
++
ωαγ0 1( ) . (2.85)
Portanto, neste modelo existe uma diferenciação entre CCIs e CCEs. A correspondência
com as curvas de sintonização neuronal é feita com o deslocamento ciliar das CCIs, ou seja,
com Di=Vi/jω: Em função da pressão transpartição teremos:
15 Notar que V0 corresponde à velocidade de deflexão dos cílios das CCEs: V0=Vr−Vt = g(VBM−Vc)−Vt. AsCCIs apresentam assim curvas de sintonização diferentes das CCEs.
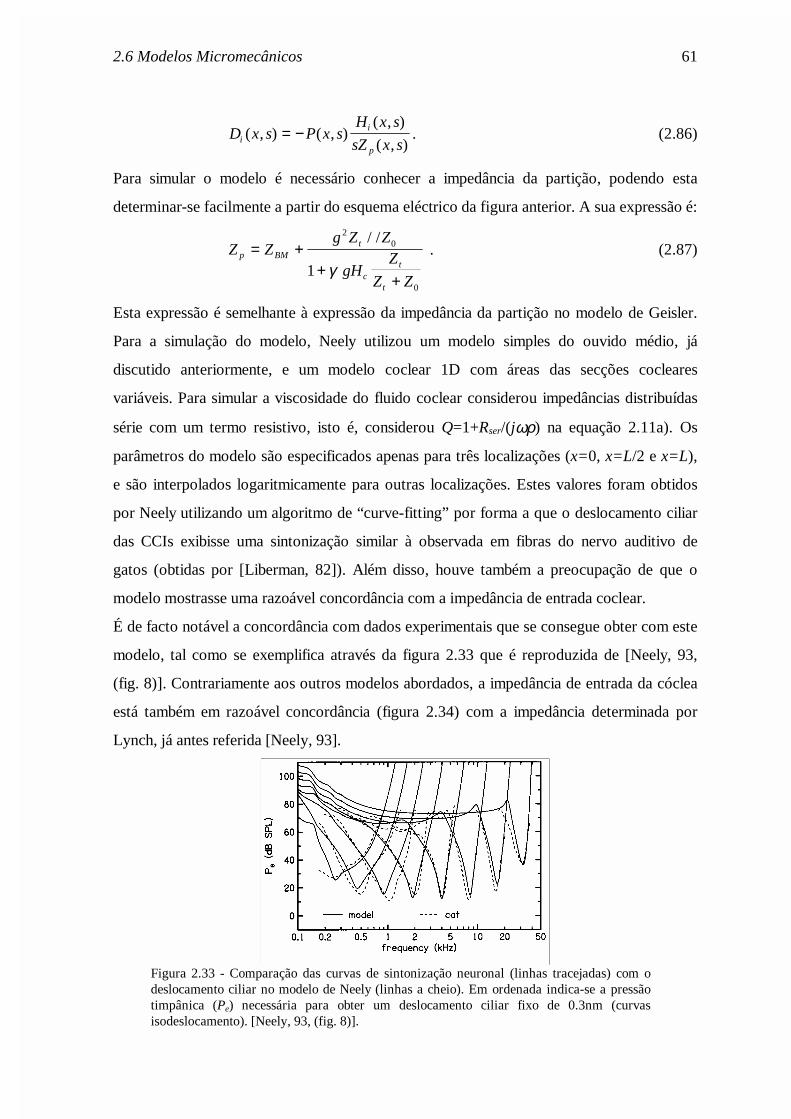
2.6 Modelos Micromecânicos 61
D x s P x sH x s
sZ x sii
p
( , ) ( , )( , )
( , )= − . (2.86)
Para simular o modelo é necessário conhecer a impedância da partição, podendo esta
determinar-se facilmente a partir do esquema eléctrico da figura anterior. A sua expressão é:
Z Zg Z Z
gHZ
Z Z
p BMt
ct
t
= ++
+
20
0
1
/ /
γ . (2.87)
Esta expressão é semelhante à expressão da impedância da partição no modelo de Geisler.
Para a simulação do modelo, Neely utilizou um modelo simples do ouvido médio, já
discutido anteriormente, e um modelo coclear 1D com áreas das secções cocleares
variáveis. Para simular a viscosidade do fluido coclear considerou impedâncias distribuídas
série com um termo resistivo, isto é, considerou Q=1+Rser/(jωρ) na equação 2.11a). Os
parâmetros do modelo são especificados apenas para três localizações (x=0, x=L/2 e x=L),
e são interpolados logaritmicamente para outras localizações. Estes valores foram obtidos
por Neely utilizando um algoritmo de “curve-fitting” por forma a que o deslocamento ciliar
das CCIs exibisse uma sintonização similar à observada em fibras do nervo auditivo de
gatos (obtidas por [Liberman, 82]). Além disso, houve também a preocupação de que o
modelo mostrasse uma razoável concordância com a impedância de entrada coclear.
É de facto notável a concordância com dados experimentais que se consegue obter com este
modelo, tal como se exemplifica através da figura 2.33 que é reproduzida de [Neely, 93,
(fig. 8)]. Contrariamente aos outros modelos abordados, a impedância de entrada da cóclea
está também em razoável concordância (figura 2.34) com a impedância determinada por
Lynch, já antes referida [Neely, 93].
Figura 2.33 - Comparação das curvas de sintonização neuronal (linhas tracejadas) com odeslocamento ciliar no modelo de Neely (linhas a cheio). Em ordenada indica-se a pressãotimpânica (Pe) necessária para obter um deslocamento ciliar fixo de 0.3nm (curvasisodeslocamento). [Neely, 93, (fig. 8)].
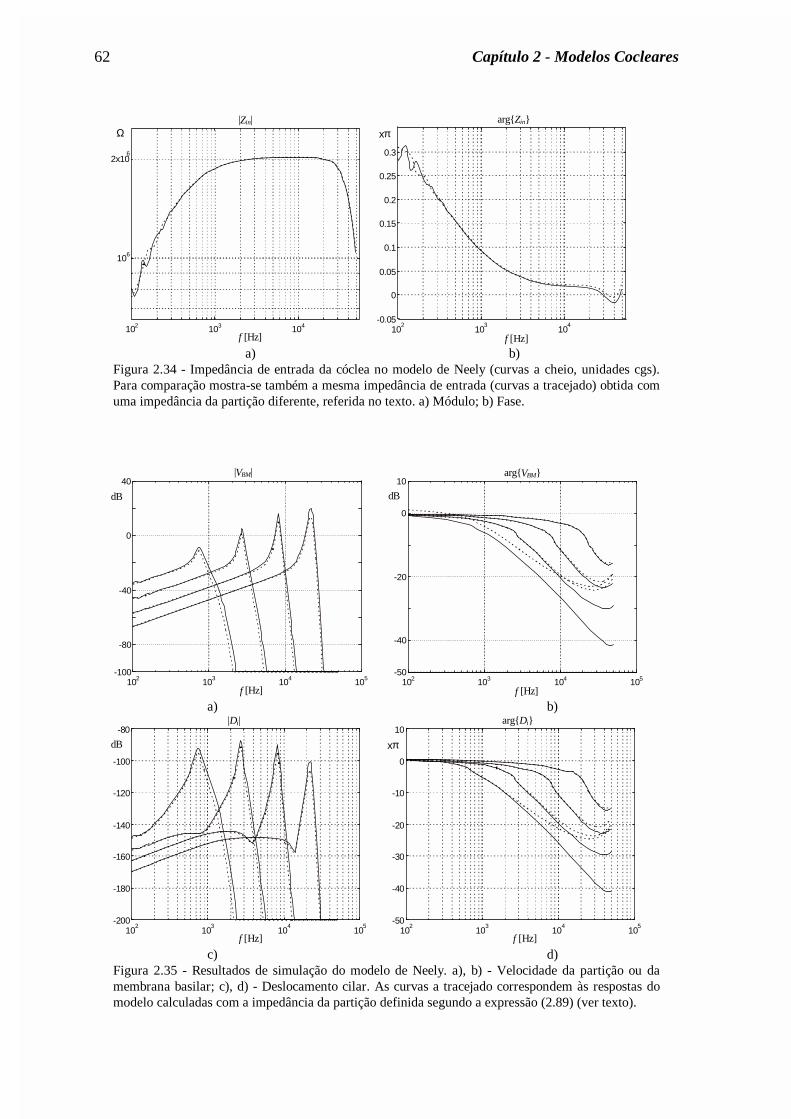
62 Capítulo 2 - Modelos Cocleares
102 103 104
106
|Zin|
f [Hz]
Ω
2x106
10
210
310
4-0.05
0
0.05
0.1
0.15
0.2
0.25
0.3
argZin
f [Hz]
xπ
a) b)Figura 2.34 - Impedância de entrada da cóclea no modelo de Neely (curvas a cheio, unidades cgs).Para comparação mostra-se também a mesma impedância de entrada (curvas a tracejado) obtida comuma impedância da partição diferente, referida no texto. a) Módulo; b) Fase.
102 103 104 105
-100
-80
-40
0
40|VBM|
f [Hz]
dB
102 103 104 105
-50
-40
-20
0
10argVBM
f [Hz]
dB
a) b)
10
210
310
410
5-200
-180
-160
-140
-120
-100
-80|Di|
f [Hz]
dB
10
210
310
410
5-50
-40
-30
-20
-10
0
10argDi
f [Hz]
xπ
c) d)Figura 2.35 - Resultados de simulação do modelo de Neely. a), b) - Velocidade da partição ou damembrana basilar; c), d) - Deslocamento cilar. As curvas a tracejado correspondem às respostas domodelo calculadas com a impedância da partição definida segundo a expressão (2.89) (ver texto).

2.6 Modelos Micromecânicos 63
Na figura 2.35 apresentam-se alguns resultados de simulação deste modelo. A velocidade
da membrana basilar não apresenta o zero espectral que caracteriza as curvas de
deslocamento ciliar, o que está essencialmente de acordo com os dados mecânicos
conhecidos.
Existem também alguns pontos fracos neste modelo. Por exemplo, a fase das respostas
apresenta uma variação demasiado rápida [Hubbard & Mountain, 96]. A massa da partição
(variável com x), apresenta valores demasiado baixos e a potência acústica injectada na
partição devido às CCEs é irrealisticamente elevada. Como se verá seguidamente, as
deficiências apontadas resultam do baixo valor da massa da membrana basilar, o que faz
com que, para as frequências de interesse, a impedância da membrana basilar no modelo
seja completamente dominada pelo termo de rigidez.
2.6.3 Análise paramétrica da impedância da partição
Como se tem vindo tornando claro a partir desta exposição, a definição da impedância da
partição joga um papel essencial na caracterização do modelo coclear. Egbert de Boer num
artigo recente [de Boer, 95], mostra a equivalência formal da impedância da partição em
alguns modelos cocleares localmente activos (essencialmente os modelos aqui revistos),
realçando a forma (2.87) ou (2.79) como definição comum. Existe contudo uma forma
alternativa de analisar a impedância da partição. Esta análise consiste em observar o
andamento das frequências naturais da impedância da partição em função da posição
coclear x e de γ. O fundamento desta análise é o seguinte: Zp deve conter uma ressonância
dominante (um par de zeros) que irá definir a frequência característica das respostas
cocleares. Além disso, Zp deve conter outros zeros e pólos de forma a reduzir ou tornar
negativa a parte real da impedância numa zona basal em relação posição de ressonância.
Assim, a impedância da partição, em vez de uma soma, pode considerar-se em termos do
produto,
Z Z Z Z Hp BM OC a a= + = , (2.88)
onde Za constitui uma impedância passiva de 2ª ordem que define a ressonância dominante
de Zp e Ha é uma função de transferência que engloba todos os outros aspectos envolvidos
na definição de Zp. Esta é uma forma alternativa e mais explícita de verificar a concordância
do modelo com os dados experimentais. Aliás, esta análise foi já feita anteriormente em
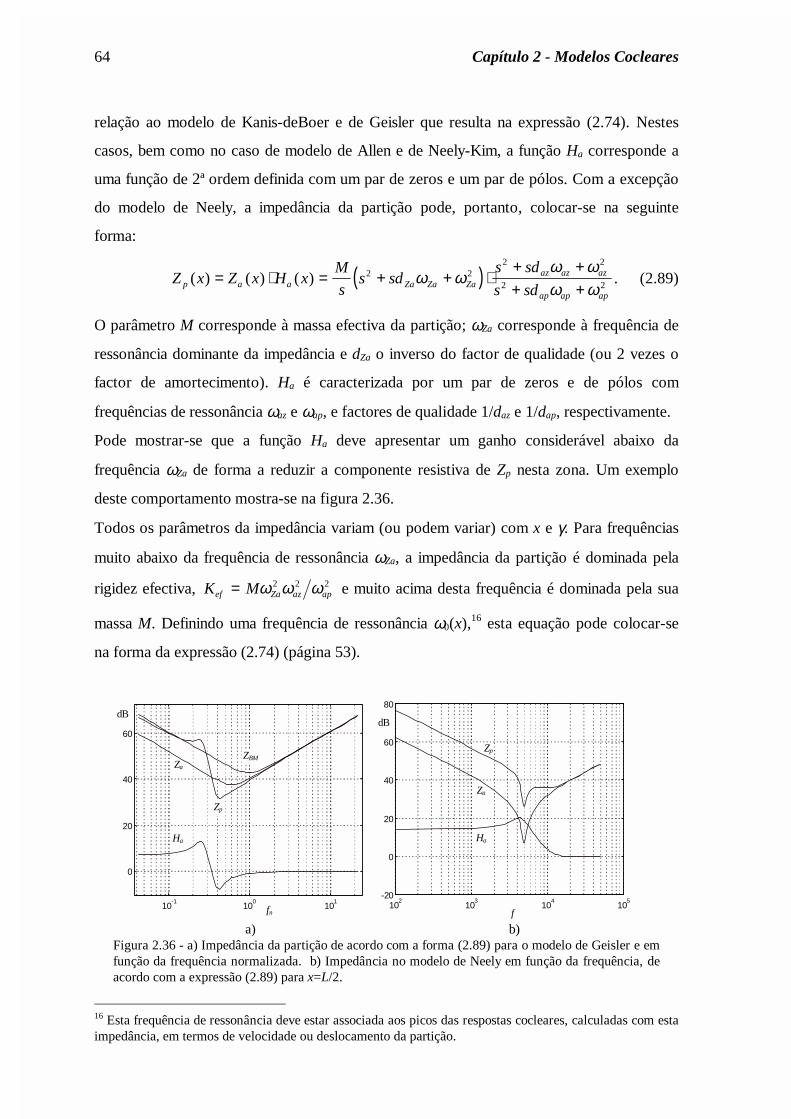
64 Capítulo 2 - Modelos Cocleares
relação ao modelo de Kanis-deBoer e de Geisler que resulta na expressão (2.74). Nestes
casos, bem como no caso de modelo de Allen e de Neely-Kim, a função Ha corresponde a
uma função de 2ª ordem definida com um par de zeros e um par de pólos. Com a excepção
do modelo de Neely, a impedância da partição pode, portanto, colocar-se na seguinte
forma:
( )Z x Z x H xM
ss sd
s sd
s sdp a a Za Za Zaaz az az
ap ap ap
( ) ( ) ( )= ⋅ = + + ⋅+ ++ +
2 22 2
2 2ω ωω ωω ω
. (2.89)
O parâmetro M corresponde à massa efectiva da partição; ωZa corresponde à frequência de
ressonância dominante da impedância e dZa o inverso do factor de qualidade (ou 2 vezes o
factor de amortecimento). Ha é caracterizada por um par de zeros e de pólos com
frequências de ressonância ωaz e ωap, e factores de qualidade 1/daz e 1/dap, respectivamente.
Pode mostrar-se que a função Ha deve apresentar um ganho considerável abaixo da
frequência ωZa de forma a reduzir a componente resistiva de Zp nesta zona. Um exemplo
deste comportamento mostra-se na figura 2.36.
Todos os parâmetros da impedância variam (ou podem variar) com x e γ. Para frequências
muito abaixo da frequência de ressonância ωZa, a impedância da partição é dominada pela
rigidez efectiva, K Mef Za az ap= ω ω ω2 2 2 e muito acima desta frequência é dominada pela sua
massa M. Definindo uma frequência de ressonância ω0(x),16 esta equação pode colocar-se
na forma da expressão (2.74) (página 53).
10
-110
010
1
0
20
40
60
dB
fn
Ha
ZBMZa
Zp
10
210
310
410
5-20
0
20
40
60
80
dB
f
Zp
Za
Ha
a) b)Figura 2.36 - a) Impedância da partição de acordo com a forma (2.89) para o modelo de Geisler e emfunção da frequência normalizada. b) Impedância no modelo de Neely em função da frequência, deacordo com a expressão (2.89) para x=L/2.
16 Esta frequência de ressonância deve estar associada aos picos das respostas cocleares, calculadas com estaimpedância, em termos de velocidade ou deslocamento da partição.

2.6 Modelos Micromecânicos 65
Analisando a impedância no modelo de Neely verifica-se que Ha tem ordem 4 e pode
colocar-se na forma seguinte:
Hs sd
s sd
s
s
s
saaz az az
ap ap ap
z
p
z
p
=+ ++ +
⋅++
⋅++
2 2
2 21
1
2
2
ω ωω ω
ωω
ωω
, (2.90)
pois constata-se que dois pólos e dois zeros são sempre reais para qualquer posição coclear.
Além disso, um zero e um pólo coincidem praticamente para qualquer x e γ (ver figura
2.38e)). Sejam eles ωz2 e ωp2. O 2º factor de 1ª ordem pode portanto suprimir-se desta
expressão. A frequência ωz1 é, neste modelo de Neely, muito superior às outras frequências
e muito superior à frequência máxima de interesse (frequência de ressonância da base da
cóclea). Assim, (s+ωz1) ≈ ωz1 e Ha decresce com a frequência e faz com que, em termos
efectivos, Zp seja dominada para altas frequências por uma resistência em vez de ser
dominada pelo termo de massa, sM. Este comportamento resulta do facto de a impedância
associada ao órgão de Corti tender para R0, o termo resistivo de Z0 e ao facto da
impedância da membrana basilar apresentar um termo de massa tão baixo que ZBM≈KB/s.
Como consequência, a constante de propagação, depois da posição de ressonância, deixa de
ser aproximadamente real o que vai provocar essencialmente uma grande variação de fase
nas respostas cocleares. Estes dois aspectos constituem uma debilidade no modelo de
Neely.
Uma forma evidente de contrariar este andamento consiste em alterar um dos zeros da
impedância, por exemplo, baixar ωz1. Fazendo coincidir ωz1 com ωp1, resulta que este factor
de 1ª ordem desaparece. Para que o ganho de Ha não se altere, deve considerar-se um factor
multiplicativo igual a ωz1/ωp1, valor este que irá assim contribuir para a um aumento
efectivo da massa da partição. Neste caso, a definição da impedância reduz-se à forma
(2.89). Na figura 2.35 as curvas a tracejado correspondem à simulação do modelo de
acordo com esta alteração. Como se pode observar, o atraso de fase máximo das respostas
cocleares reduz-se para cerca de metade, sendo mesmo assim, ainda demasiado acentuado
tendo em conta os valores indicados por Hubbard e Mountain, [Hubbard & Mountain, 96].
O módulo das respostas reduz-se apenas moderadamente.
De salientar que esta alteração da impedância da partição apenas é utilizada na
determinação da velocidade da partição e não na determinação da velocidade ou
deslocamento ciliar.
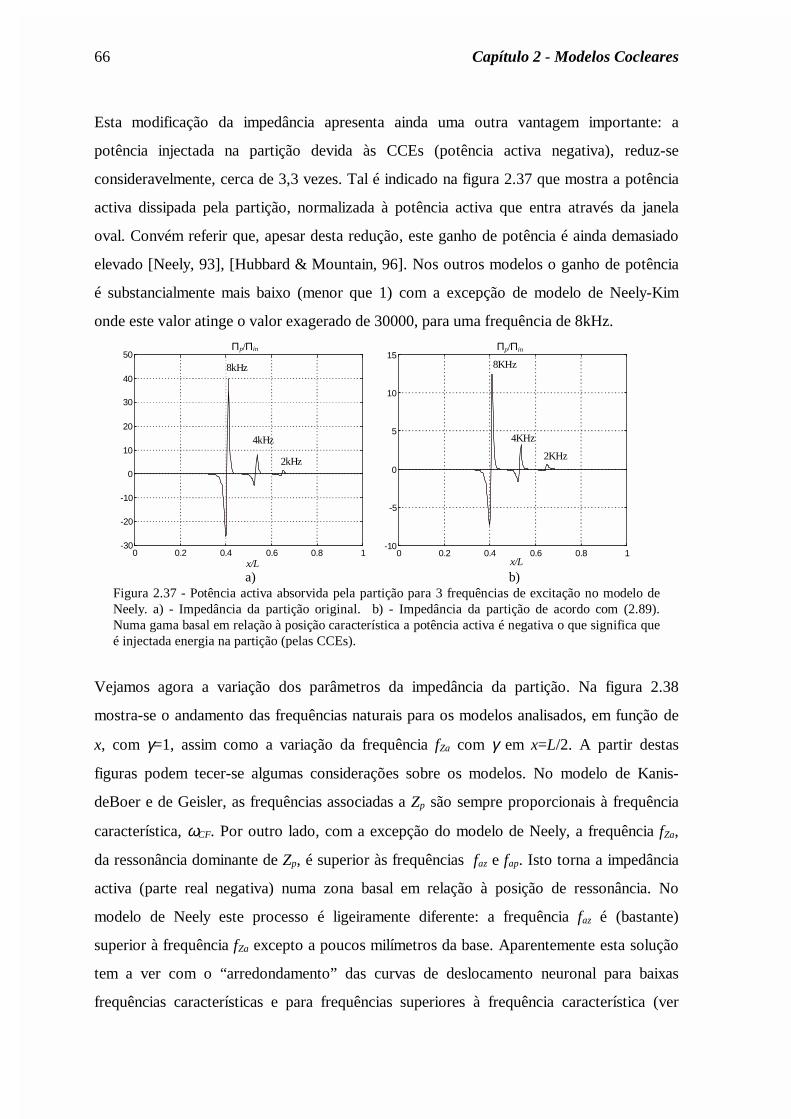
66 Capítulo 2 - Modelos Cocleares
Esta modificação da impedância apresenta ainda uma outra vantagem importante: a
potência injectada na partição devida às CCEs (potência activa negativa), reduz-se
consideravelmente, cerca de 3,3 vezes. Tal é indicado na figura 2.37 que mostra a potência
activa dissipada pela partição, normalizada à potência activa que entra através da janela
oval. Convém referir que, apesar desta redução, este ganho de potência é ainda demasiado
elevado [Neely, 93], [Hubbard & Mountain, 96]. Nos outros modelos o ganho de potência
é substancialmente mais baixo (menor que 1) com a excepção de modelo de Neely-Kim
onde este valor atinge o valor exagerado de 30000, para uma frequência de 8kHz.
0 0.2 0.4 0.6 0.8 1
-30
-20
-10
0
10
20
30
40
50
x/L
Πp/Πin
8kHz
4kHz
2kHz
0 0.2 0.4 0.6 0.8 1
-10
-5
0
5
10
15
x/L
8KHz
4KHz
2KHz
Πp/Πin
a) b)Figura 2.37 - Potência activa absorvida pela partição para 3 frequências de excitação no modelo deNeely. a) - Impedância da partição original. b) - Impedância da partição de acordo com (2.89).Numa gama basal em relação à posição característica a potência activa é negativa o que significa queé injectada energia na partição (pelas CCEs).
Vejamos agora a variação dos parâmetros da impedância da partição. Na figura 2.38
mostra-se o andamento das frequências naturais para os modelos analisados, em função de
x, com γ=1, assim como a variação da frequência fZa com γ em x=L/2. A partir destas
figuras podem tecer-se algumas considerações sobre os modelos. No modelo de Kanis-
deBoer e de Geisler, as frequências associadas a Zp são sempre proporcionais à frequência
característica, ωCF. Por outro lado, com a excepção do modelo de Neely, a frequência fZa,
da ressonância dominante de Zp, é superior às frequências faz e fap. Isto torna a impedância
activa (parte real negativa) numa zona basal em relação à posição de ressonância. No
modelo de Neely este processo é ligeiramente diferente: a frequência faz é (bastante)
superior à frequência fZa excepto a poucos milímetros da base. Aparentemente esta solução
tem a ver com o “arredondamento” das curvas de deslocamento neuronal para baixas
frequências características e para frequências superiores à frequência característica (ver
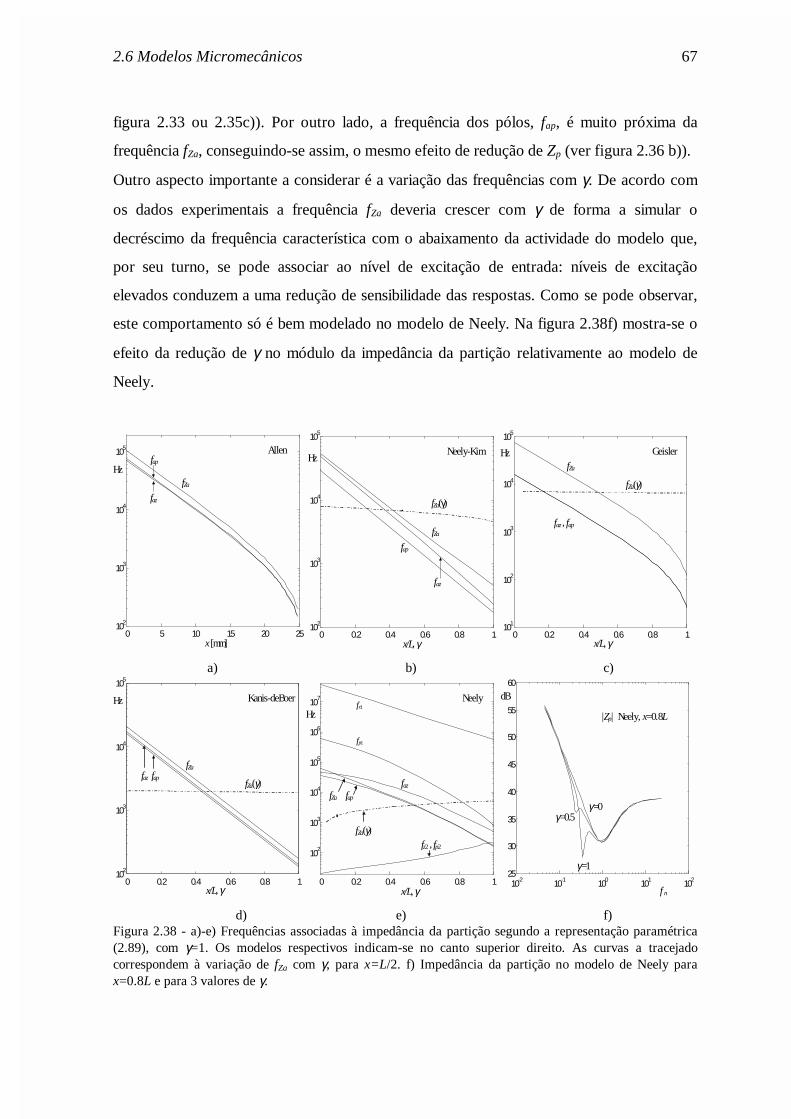
2.6 Modelos Micromecânicos 67
figura 2.33 ou 2.35c)). Por outro lado, a frequência dos pólos, fap, é muito próxima da
frequência fZa, conseguindo-se assim, o mesmo efeito de redução de Zp (ver figura 2.36 b)).
Outro aspecto importante a considerar é a variação das frequências com γ. De acordo com
os dados experimentais a frequência fZa deveria crescer com γ de forma a simular o
decréscimo da frequência característica com o abaixamento da actividade do modelo que,
por seu turno, se pode associar ao nível de excitação de entrada: níveis de excitação
elevados conduzem a uma redução de sensibilidade das respostas. Como se pode observar,
este comportamento só é bem modelado no modelo de Neely. Na figura 2.38f) mostra-se o
efeito da redução de γ no módulo da impedância da partição relativamente ao modelo de
Neely.
0 5 10 15 20 25102
103
104
105
fZa
x [mm]
Hz
faz
fap
Allen
0 0.2 0.4 0.6 0.8 1102
103
104
105
Hz
fZa
fap
faz
fZa(γ)
x/L, γ
Neely-Kim
0 0.2 0.4 0.6 0.8 1101
102
103
104
105
x/L, γ
Hz fZa
faz , fap
fZa(γ)
Geisler
a) b) c)
0 0.2 0.4 0.6 0.8 110
2
103
104
105
x/L, γ
Hz
fZa
faz fap fZa(γ)
Kanis-deBoer
0 0.2 0.4 0.6 0.8 1
102
103
104
105
106
107
x/L, γ
Hz
fZa
faz
fap
fz2 , fp2
fz1
fp1
fZa(γ)
Neely
10-2 10-1 100 101 10225
30
35
40
45
50
55
60
dB
f n
|Zp| Neely, x=0.8L
γ =1
γ =0.5 γ =0
d) e) f)Figura 2.38 - a)-e) Frequências associadas à impedância da partição segundo a representação paramétrica(2.89), com γ=1. Os modelos respectivos indicam-se no canto superior direito. As curvas a tracejadocorrespondem à variação de fZa com γ, para x=L/2. f) Impedância da partição no modelo de Neely parax=0.8L e para 3 valores de γ.

68 Capítulo 2 - Modelos Cocleares
2.6.4 Outros modelos
Os modelos analisados na secção 2.6.2 representam o estado da arte dos desenvolvimentos
mais recentes da modelação coclear. Existem contudo, ainda alguns modelos cocleares que
merecem referência. É o caso do modelo de Mammano e Nobili [Mammano & Nobili, 93]
que parte de uma formulação idêntica à descrita anteriormente, mas difere quanto à maneira
como as membranas basilar e tectória se movem. O modelo é descrito por uma equação
integral que utiliza a função de Green, já utilizada por Sondhi [Sondhi, 78] na sua
formulação 2D. Os resultados obtidos com este modelo são concordantes com os dados da
velocidade da membrana basilar [Hubbard & Mountain, 96]. Um outro modelo importante é
o modelo proposto por Neely e Stover [Neely & Stover, 1993]. Trata-se de um modelo
essencialmente idêntico aos modelos de Geisler e de Neely, com a mesma formulação, mas
onde uma ressonância é incluída no órgão de Corti. As respostas de deslocamento ciliar
apresentam uma relação de amplitude pico-cauda bastante inferior às conseguidas com o
modelo de Neely. O modelo é usado para estudar emissões otoacústicas e é proposta uma
versão não linear do modelo, no domínio temporal, onde o parâmetro γ se toma
inversamente proporcional ao módulo do deslocamento ciliar. Geisler propôs também
recentemente [Geisler & Sang, 95] um outro modelo, ainda com a mesma formulação que o
anteriormente descrito, mas onde as CCEs estão dispostas com uma dada inclinação
longitudinal. Desta forma, uma simples CCE não pertence a uma única secção coclear
podendo produzir forças em diferentes posições longitudinais, aliás como tinha já sido
proposto por Steele [Steele et al, 93]. As respostas obtidas não parecem tão concordantes
com os dados experimentais como as dos modelo anteriores.
Modelos com uma formulação diferente foram também propostos. Tratam-se de modelos
denominados multimodo, que admitem que o órgão de Corti ou as estruturas associadas
têm a capacidade de propagar ondas. É o caso do modelo “sandwich” [de Boer, 91] que
admite a existência de um canal adicional no interior do órgão de Corti onde ondas
acústicas se podem propagar. Outro modelo deste tipo é o modelo de dupla linha de
transmissão proposto por Hubbard [Hubbard, 93], que admite dois modos de propagação
separados. O acoplamento entre as duas linhas de transmissão é feito por elementos activos
que representam as CCEs. Apesar de se tratar de um modelo fenomenológico este modelo
apresenta respostas bastante concordantes com os dados experimentais.

2.7 Modelo da Cóclea Humana 69
Existem ainda modelos cocleares que utilizam técnicas de processamento de sinal,
nomeadamente bancos de filtros, para simular as respostas cocleares, sendo este assunto
discutido no capítulo seguinte. Com base neste conceito foram desenvolvidos circuitos
integrados que simulam a cóclea [Lyon & Mead, 88], [Lazzaro, 91], [Watts et al, 92], [Liu
et al, 92], [Lazzaro et al, 93], [Bor & Wu, 96] e que podem ser utilizados como módulos de
pré-processamento para análise de sinais audio com interesse em reconhecimento
automático da fala [Fragnière, 96].
Muito recentemente surgiram trabalhos onde se utiliza a técnica dos elementos finitos para
descrever a cóclea a três dimensões [Kolston & Ashmore, 96], [Funnell, 96]. A investigação
recente sobre modelação coclear tem-se centrado ultimamente na adequação dos modelos
na explicação de fenómenos não lineares na interacção de par de tons: geração de tons de
distorção (DP - “distortion products”) [Kanis & de Boer, 97] e supressão por dois tons.
Algumas das questões ainda não esclarecidas sobre a micromecânica da cóclea poderão vir
a ser resolvidas num futuro próximo, à medida que novos dados experimentais forem
surgindo.
2.7 Modelo da Cóclea Humana
No capítulo seguinte será explorada a possibilidade de utilizar um modelo coclear num
esquema de banco de filtros. Para o processamento de fala interessa considerar um modelo
da cóclea humana. Os modelos antes abordados tentam reproduzir os dados fisiológicos
existentes e são por isso, na maior parte dos casos, definidos para a cóclea do gato ou
outros mamíferos de pequeno porte. Aborda-se nesta secção os problemas envolvidos na
conversão de um modelo coclear, tal como definido originalmente, para a cóclea humana.
Pretende-se reter a definição da impedância da partição, fazendo apenas pequenas
alterações que conduzam basicamente a uma variação das frequências características que
corresponda agora à cóclea humana. As diferenças mais significativas a ter em conta são as
seguintes:
a) a máxima frequência detectável, ou seja a frequência de ressonância na base da cóclea, é
para o ouvido humano cerca de 20kHz em vez dos cerca de 50kHz do gato;

70 Capítulo 2 - Modelos Cocleares
b) a frequência mais baixa, no ápice, é apenas ligeiramente mais baixa (cerca de 20-30Hz
em vez de cerca de 100Hz do gato);
c) a cóclea humana é mais longa, (cerca de 3,5 cm em vez de 2,5 cm do gato) e com áreas
das secções cocleares maiores. Isto faz com que as impedâncias acústicas da entrada da
cóclea e do ouvido médio sejam inferiores.
Para a cóclea humana não existem dados fisiológicos de interesse a não ser dados obtidos
em cadáveres [Békésy, 60], [Puria et al, 93]. Existem contudo dados psicofísicos a partir
dos quais se podem inferir algumas propriedades da cóclea. Por outro lado, o estudo
comparativo de várias espécies de mamíferos [Greenwood, 90], indica que entre elas existe
uma perfeita analogia quanto ao funcionamento do sistema auditivo periférico.
Uma experiência psicofísica, em torno da qual existe muita investigação, corresponde à
obtenção de curvas de sintonização (PTCs - “Psychophysical Tuning Curves”) em analogia
com as curvas de sintonização neuronal [Zwicker, 74], [Moore, 78], [Davies & Patterson,
79], [Glasberg et al, 84], [Nelson et al, 90], [Delgute, 90]. Em particular, um paradigma
proposto por Zwicker [Zwicker, 74], em perfeita analogia com o método de determinação
de sintonia neuronal, consiste em determinar qual a intensidade de um tom de máscara
necessário para ser efectivo a mascarar um tom de teste, de baixa intensidade, perto do
limiar auditivo e frequência fixa. As curvas obtidas, (figura 2.39) são, de forma
impressionante, parecidas com as curvas de sintonia neuronal e sugerem que a explicação
do fenómeno de mascaramento (“masking”) é basicamente determinado pelo mecanismo
auditivo periférico e que não necessita de maior elaboração em centros nervosos superiores
para o explicar [Buser & Imbert, 92].
Figura 2.39 - Limiares de mascaramento para 3 tons de teste de 630 Hz, 2 kHz e 8 kHz (curvas daesquerda para a direita), com nível de pressão sonora 5dB acima do limiar absoluto (x). O nívelde pressão sonora, LM, é o nível do tom de máscara que é efectivo para mascarar o som de testeque tem frequência e intensidade fixa (in [Zwicker, 74]).
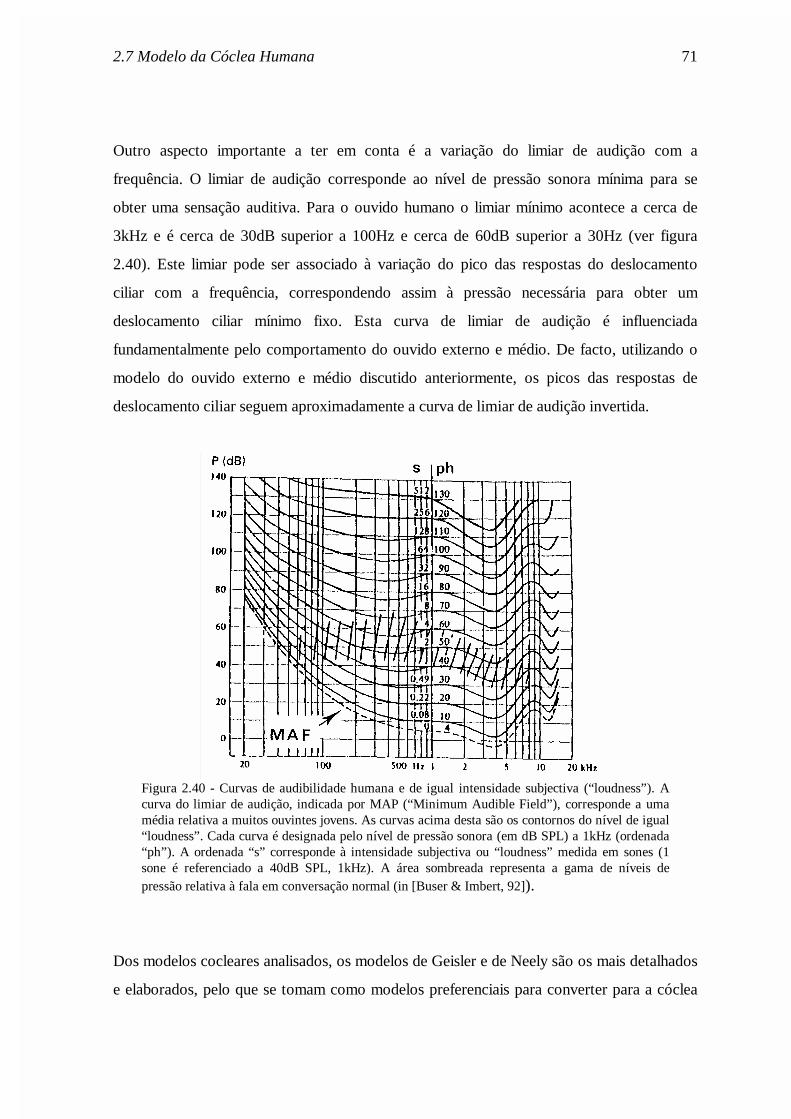
2.7 Modelo da Cóclea Humana 71
Outro aspecto importante a ter em conta é a variação do limiar de audição com a
frequência. O limiar de audição corresponde ao nível de pressão sonora mínima para se
obter uma sensação auditiva. Para o ouvido humano o limiar mínimo acontece a cerca de
3kHz e é cerca de 30dB superior a 100Hz e cerca de 60dB superior a 30Hz (ver figura
2.40). Este limiar pode ser associado à variação do pico das respostas do deslocamento
ciliar com a frequência, correspondendo assim à pressão necessária para obter um
deslocamento ciliar mínimo fixo. Esta curva de limiar de audição é influenciada
fundamentalmente pelo comportamento do ouvido externo e médio. De facto, utilizando o
modelo do ouvido externo e médio discutido anteriormente, os picos das respostas de
deslocamento ciliar seguem aproximadamente a curva de limiar de audição invertida.
Figura 2.40 - Curvas de audibilidade humana e de igual intensidade subjectiva (“loudness”). Acurva do limiar de audição, indicada por MAP (“Minimum Audible Field”), corresponde a umamédia relativa a muitos ouvintes jovens. As curvas acima desta são os contornos do nível de igual“loudness”. Cada curva é designada pelo nível de pressão sonora (em dB SPL) a 1kHz (ordenada“ph”). A ordenada “s” corresponde à intensidade subjectiva ou “loudness” medida em sones (1sone é referenciado a 40dB SPL, 1kHz). A área sombreada representa a gama de níveis depressão relativa à fala em conversação normal (in [Buser & Imbert, 92]).
Dos modelos cocleares analisados, os modelos de Geisler e de Neely são os mais detalhados
e elaborados, pelo que se tomam como modelos preferenciais para converter para a cóclea
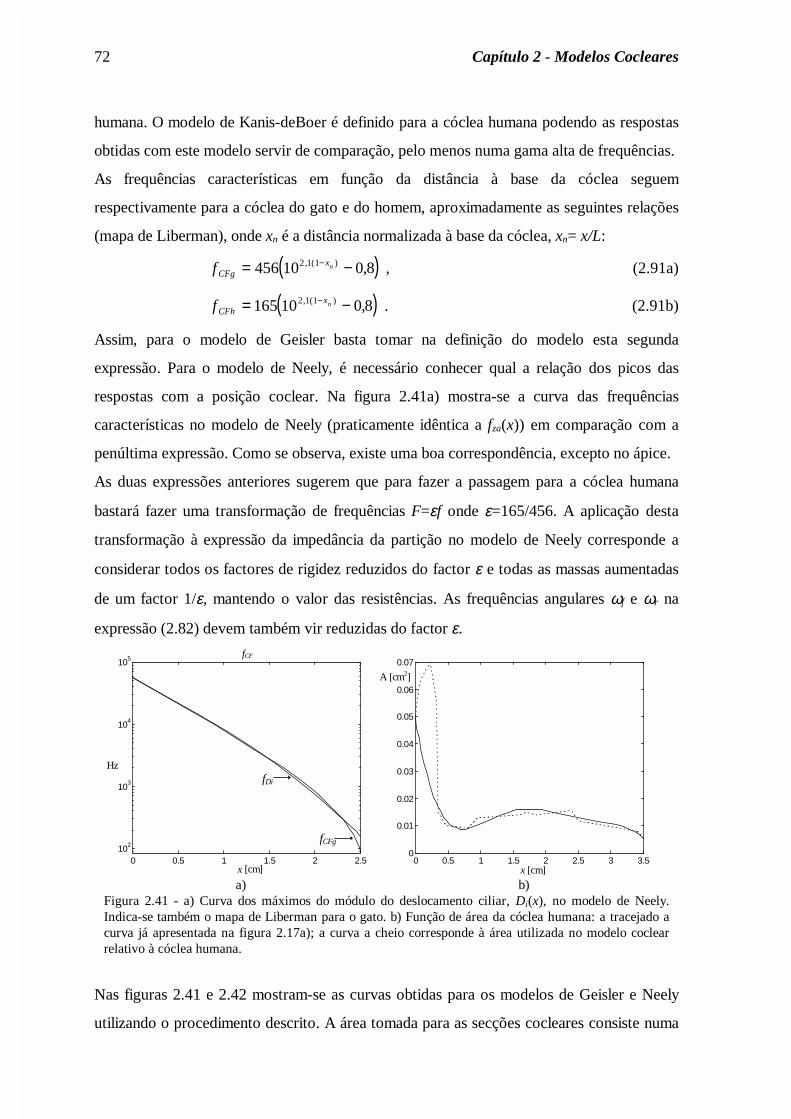
72 Capítulo 2 - Modelos Cocleares
humana. O modelo de Kanis-deBoer é definido para a cóclea humana podendo as respostas
obtidas com este modelo servir de comparação, pelo menos numa gama alta de frequências.
As frequências características em função da distância à base da cóclea seguem
respectivamente para a cóclea do gato e do homem, aproximadamente as seguintes relações
(mapa de Liberman), onde xn é a distância normalizada à base da cóclea, xn= x/L:
( )f CFgxn= −−456 10 0 82 1 1, ( ) , , (2.91a)
( )f CFhxn= −−165 10 0 82 1 1, ( ) , . (2.91b)
Assim, para o modelo de Geisler basta tomar na definição do modelo esta segunda
expressão. Para o modelo de Neely, é necessário conhecer qual a relação dos picos das
respostas com a posição coclear. Na figura 2.41a) mostra-se a curva das frequências
características no modelo de Neely (praticamente idêntica a fza(x)) em comparação com a
penúltima expressão. Como se observa, existe uma boa correspondência, excepto no ápice.
As duas expressões anteriores sugerem que para fazer a passagem para a cóclea humana
bastará fazer uma transformação de frequências F=εf onde ε=165/456. A aplicação desta
transformação à expressão da impedância da partição no modelo de Neely corresponde a
considerar todos os factores de rigidez reduzidos do factor ε e todas as massas aumentadas
de um factor 1/ε, mantendo o valor das resistências. As frequências angulares ωf e ωr na
expressão (2.82) devem também vir reduzidas do factor ε.
0 0.5 1 1.5 2 2.5
102
103
104
105 fCF
x [cm]
Hz
fCFg
fDi
0 0.5 1 1.5 2 2.5 3 3.5
0
0.01
0.02
0.03
0.04
0.05
0.06
0.07
x [cm]
A [cm2]
a) b)Figura 2.41 - a) Curva dos máximos do módulo do deslocamento ciliar, Di(x), no modelo de Neely.Indica-se também o mapa de Liberman para o gato. b) Função de área da cóclea humana: a tracejado acurva já apresentada na figura 2.17a); a curva a cheio corresponde à área utilizada no modelo coclearrelativo à cóclea humana.
Nas figuras 2.41 e 2.42 mostram-se as curvas obtidas para os modelos de Geisler e Neely
utilizando o procedimento descrito. A área tomada para as secções cocleares consiste numa
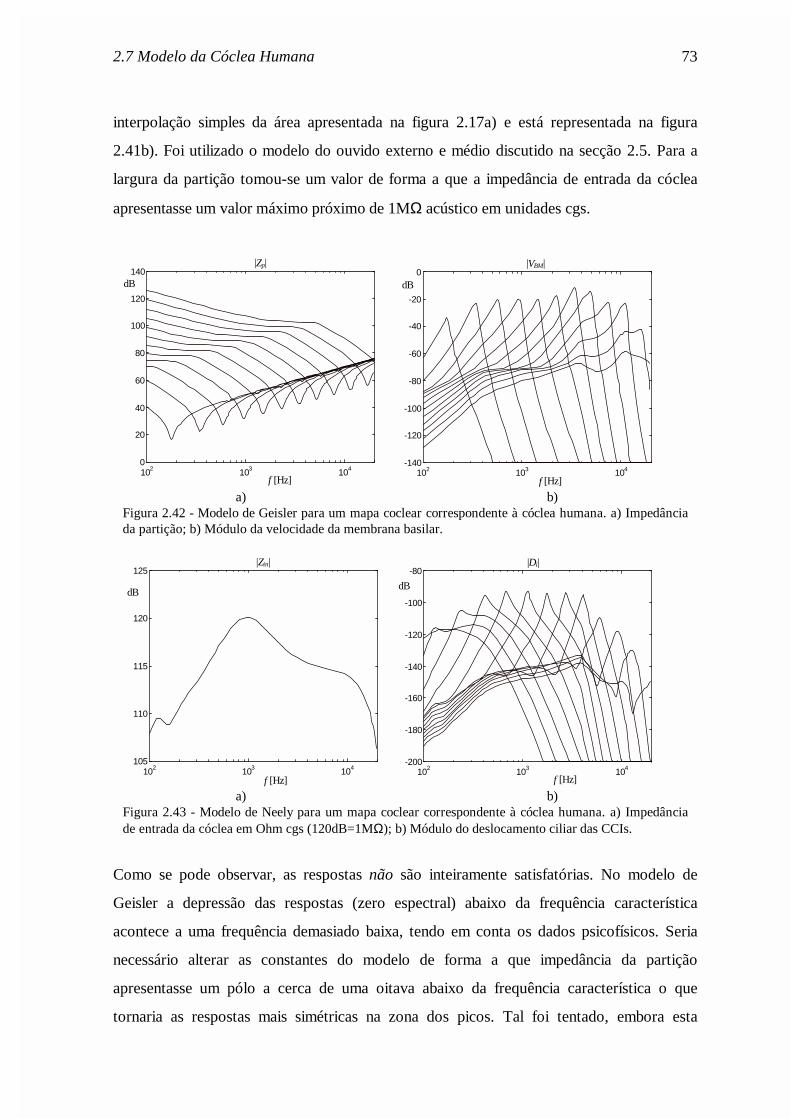
2.7 Modelo da Cóclea Humana 73
interpolação simples da área apresentada na figura 2.17a) e está representada na figura
2.41b). Foi utilizado o modelo do ouvido externo e médio discutido na secção 2.5. Para a
largura da partição tomou-se um valor de forma a que a impedância de entrada da cóclea
apresentasse um valor máximo próximo de 1MΩ acústico em unidades cgs.
102 103 1040
20
40
60
80
100
120
140|Zp|
f [Hz]
dB
102 103 104-140
-120
-100
-80
-60
-40
-20
0|VBM|
f [Hz]
dB
a) b)Figura 2.42 - Modelo de Geisler para um mapa coclear correspondente à cóclea humana. a) Impedânciada partição; b) Módulo da velocidade da membrana basilar.
102 103 104
105
110
115
120
125|Zin|
f [Hz]
dB
102 103 104
-200
-180
-160
-140
-120
-100
-80|Di|
f [Hz]
dB
a) b)Figura 2.43 - Modelo de Neely para um mapa coclear correspondente à cóclea humana. a) Impedânciade entrada da cóclea em Ohm cgs (120dB=1MΩ); b) Módulo do deslocamento ciliar das CCIs.
Como se pode observar, as respostas não são inteiramente satisfatórias. No modelo de
Geisler a depressão das respostas (zero espectral) abaixo da frequência característica
acontece a uma frequência demasiado baixa, tendo em conta os dados psicofísicos. Seria
necessário alterar as constantes do modelo de forma a que impedância da partição
apresentasse um pólo a cerca de uma oitava abaixo da frequência característica o que
tornaria as respostas mais simétricas na zona dos picos. Tal foi tentado, embora esta

74 Capítulo 2 - Modelos Cocleares
mudança produza um efeito indesejado: o abaixamento da frequência dos pólos de Zp faz
com que a relação pico-cauda das respostas seja demasiado baixa. Por outro lado,
aumentando o factor de qualidade dominante na impedância Zp produzem-se picos muito
finos tal como acontece no modelo de Kanis-deBoer. A existência de uma filtragem
adicional relativamente às CCIs, tal como é proposto no modelo de Neely pode resolver
esta questão. Uma solução satisfatória é apresentada em [Perdigão et al, 96] onde se
considera a impedância da partição definida parametricamente. Neste caso, a relação pico-
cauda das respostas é cerca de 30 dB, portanto, um pouco baixa.
Quanto ao modelo de Neely, as respostas parecem mais condizentes com as curvas de
sintonia psicofísica (PTCs) e o limiar de audição, correspondente ao andamento na
frequência dos picos das respostas, é razoavelmente representado. A impedância de entrada
da cóclea é também muito condizente com os resultados preditos por Puria e Allen [Puria &
Allen, 91] (figura 2.43a)). Contudo, as depressões nas respostas correspondentes ao zero
espectral parecem demasiado acentuadas e as curvas correspondentes a baixas CFs têm
larguras de banda demasiado elevadas. Além disso, a relação de amplitudes pico-cauda das
respostas é demasiado elevada (cerca de 60dB). Foram feitas várias tentativas de alteração
dos parâmetros físicos (resistências massas e factores de rigidez) da impedância da partição
de forma a conciliar estes aspectos mas sem sucesso.
Uma descrição paramétrica simples da impedância da partição, tal como é apresentada na
secção anterior, pode conduzir à definição de modelos com respostas mais razoáveis. Tal é
mostrado em [Perdigão et al, 96] onde a definição da impedância da partição é tomada de
acordo com o modelo de Geisler. O mesmo tipo de modelação foi também aplicado ao
modelo de Neely. A impedância da partição é definida tal como em (2.89) onde a frequência
fZa em função de x, corresponde ao mapa coclear (2.91b). A frequência faz é superior a fZa e
fap ligeiramente inferior, tal como no modelo de Neely original17. A massa da partição
coclear é agora tomada constante, com um valor razoável, tal como no modelo de Kanis-
deBoer. Aplica-se seguidamente uma filtragem adicional ao deslocamento da membrana
basilar por uma função Hi de forma a obter o deslocamento ciliar relativo às CCIs, tal como
é proposto por Neely (expressão (2.86)). Hi apresenta um par de zeros uma oitava abaixo
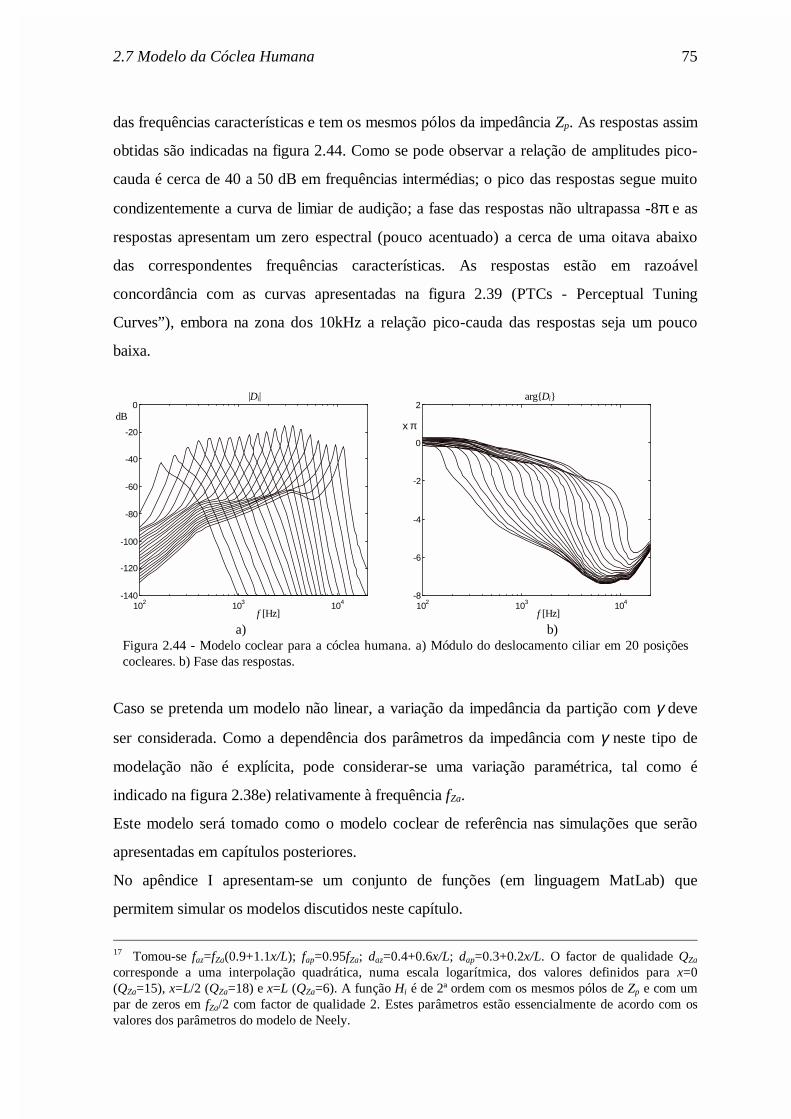
2.7 Modelo da Cóclea Humana 75
das frequências características e tem os mesmos pólos da impedância Zp. As respostas assim
obtidas são indicadas na figura 2.44. Como se pode observar a relação de amplitudes pico-
cauda é cerca de 40 a 50 dB em frequências intermédias; o pico das respostas segue muito
condizentemente a curva de limiar de audição; a fase das respostas não ultrapassa -8π e as
respostas apresentam um zero espectral (pouco acentuado) a cerca de uma oitava abaixo
das correspondentes frequências características. As respostas estão em razoável
concordância com as curvas apresentadas na figura 2.39 (PTCs - Perceptual Tuning
Curves”), embora na zona dos 10kHz a relação pico-cauda das respostas seja um pouco
baixa.
102 103 104-140
-120
-100
-80
-60
-40
-20
0|Di|
f [Hz]
dB
102 103 104-8
-6
-4
-2
0
2
f [Hz]
x π
argDi
a) b)Figura 2.44 - Modelo coclear para a cóclea humana. a) Módulo do deslocamento ciliar em 20 posiçõescocleares. b) Fase das respostas.
Caso se pretenda um modelo não linear, a variação da impedância da partição com γ deve
ser considerada. Como a dependência dos parâmetros da impedância com γ neste tipo de
modelação não é explícita, pode considerar-se uma variação paramétrica, tal como é
indicado na figura 2.38e) relativamente à frequência fZa.
Este modelo será tomado como o modelo coclear de referência nas simulações que serão
apresentadas em capítulos posteriores.
No apêndice I apresentam-se um conjunto de funções (em linguagem MatLab) que
permitem simular os modelos discutidos neste capítulo.
17 Tomou-se faz=fZa(0.9+1.1x/L); fap=0.95fZa; daz=0.4+0.6x/L; dap=0.3+0.2x/L. O factor de qualidade QZa
corresponde a uma interpolação quadrática, numa escala logarítmica, dos valores definidos para x=0(QZa=15), x=L/2 (QZa=18) e x=L (QZa=6). A função Hi é de 2ª ordem com os mesmos pólos de Zp e com umpar de zeros em fZa/2 com factor de qualidade 2. Estes parâmetros estão essencialmente de acordo com osvalores dos parâmetros do modelo de Neely.

76 Capítulo 2 - Modelos Cocleares
2.8 Conclusões
Apesar de existirem ainda um conjunto de hipóteses competitivas sobre o funcionamento
micromecânico da cóclea, pode dizer-se que a modelação coclear atingiu um estado de
maturação em que as respostas cocleares podem ser simuladas com razoável detalhe.
Os modelos activos revistos incluem uma série de funcionalidades que cuja existência tem
vindo a ser comprovada. Além disso, conduzem a resultados concordantes com os dados
obtidos através de preparações experimentais em cócleas vivas. Nomeadamente, os modelos
consideram elementos activos associados às CCEs; incluem uma realimentação negativa que
influencia a vibração da partição coclear amplificando a onda progressiva na cóclea e
incluem um segundo filtro com um zero espectral proeminente em concordância com os
dados neuronais. De entre os modelos revistos o modelo de Neely e de Geisler são os mais
completos e concordantes com dados conhecidos.
A maior parte dos modelos desenvolvidos tenta simular dados mecânicos obtidos em
mamíferos (principalmente gatos, porcos da Guiné e chinchilas). Dados relativos à cóclea
humana são escassos e, por razões óbvias, nunca são feitas experiências em cócleas
humanas vivas. Porém, existe uma clara evidência que o aparelho auditivo periférico é
perfeitamente semelhante e análogo entre os mamíferos [Greenwood, 90].
A conversão dos modelos para a simulação da cóclea humana, apesar de parecer um tarefa
simples e directa, apresenta alguns problemas difíceis de solucionar. No entanto, foi
apresentado um modelo que se pode considerar suficientemente preciso tendo em conta os
dados conhecidos. A definição deste modelo resulta de uma parametrização da impedância
da partição, diferente da que é usual considerar, mas com a qual é possível controlar de uma
forma mais explícita as características das respostas cocleares. Tanto quanto é possível
avaliar pela literatura especializada, não existem publicados modelos relativos à cóclea
humana com respostas tão plausíveis quanto as apresentadas pelo modelo proposto.
Este modelo será utilizado no desenvolvimento de um modelo discreto no tempo, baseado
em bancos de filtros, a ser especificado no capítulo seguinte.


















