
Biologia wzrostu i rozwoju dwu odmian Dictamnus albus L ... · nego wykazały, że zachowanie tego...
16
21 Postępy Fitoterapii 1/2015 Borgis Post Fitoter 2015; (16)1: 21-36 Prezentowana praca jest kontynuacją badań nad systematyką gatunku Dictamnus, jego rozmieszcze- niem w Europie i na świecie oraz przeglądem ważniej- szych grup związków biologicznie czynnych, ich bio- genezy, właściwości, występowania i zastosowania (1). W pierwszej części opisano alkaloidy furochinolinowe, furanokumaryny i limonoidy, zaś w drugiej omówiono olejki eteryczne, flawonoidy oraz inne związki biolo- gicznie aktywne (2, 3). Dyptam jesionolistny znany jest pod nazwą „go- rejącego krzewu Mojżesza”. Nazwa ta pochodzi od opisu biblijnego rośliny: „Wtedy ukazał mu się Anioł Pański w płomieniu ognia, ze środka krzewu. Moj- żesz widział, jak krzew płonął ogniem, a nie spłonął od niego.” (Biblia, Księga Wyjścia). Dziś wiemy, że zjawisko to było możliwe gdyż wszystkie struktury w kwiatostanach dyptamu są gęsto pokryte gruczo- łami wydzielniczymi. Produkowane przez nie obficie olejki eteryczne w upalne, bezwietrzne dni mogą ulec samozapłonowi, nie uszkadzając rośliny (4, 5). Postępujące skażenie środowiska naturalnego jest przyczyną zamierania wielu gatunków roślin, rosną- cych nawet na terenach parków narodowych i rezer- watów przyrody. Do takich ginących roślin należy zaliczyć Dictamnus albus L. (dyptam jesionolistny). Jako roślina reliktowa i bardzo rzadka, zawierająca ponadto szereg osobliwości strukturalnych i biologicz- nych, podlega ona całkowitej ochronie. Istnieje więc konieczność czynnej ochrony najbardziej zagrożonych populacji tych roślin w kontrolowanych warunkach. Mnożenie generatywne i uprawa dyptamu jesionolist- nego wykazały, że zachowanie tego gatunku w warun- kach ex situ daje pozytywne rezultaty. Dlatego też jest ona coraz częściej uprawiana jako roślina ozdobna o wyjątkowych walorach dekoracyjnych (6, 7). *Tadeusz Wolski 1,2 , Tomasz Baj 1 Biologia wzrostu i rozwoju dwu odmian dyptamu jesionolistnego (Dictamnus albus L. cv. Albiflores i cv. Rosa Purple) oraz budowa anatomiczno-morfologiczna struktur wydzielniczych, skład olejku eterycznego i analiza ekstraktu 1 Katedra i Zakład Farmakognozji z Pracownią Roślin Leczniczych, Uniwersytet Medyczny w Lublinie Kierownik Katedry i Zakładu: dr hab. Grażyna Zgórka 2 Katedra Warzywnictwa i Roślin Leczniczych, Uniwersytet Przyrodniczy w Lublinie Kierownik Katedry: prof. dr hab. Jan Dyduch BIOLOGY OF GROWTH AND DEVELOPMENT OF TWO CULTIVARS OF DICTAMNUS ALBUS L. (CV. ALBIFLORES AND CV. ROSA PURPLE), ANATOMICAL AND MORPHOLOGICAL STRUCTURE OF SECRETORY TISSUES AND ANALYSIS OF THE ESSENTIAL OIL SUMMARY The paper presents the biology of growth and development of two varieties of dittany (Dictamnus albus L. cv. Albiflores and cv. Rosa Purple). The present study include biometric measurements of morphological characteristics, including the flowering and pollination dynamics and efficiency of pollen. Measurements of the amount of shoots, stems of length and a width and length of the leaves. Set the number of flowering shoots, as well as the number of flowers on the shoot and the number of flowers per plant, given the dimensions of the grains of pollen, nectar and mass concentrations of sugar and honey yield. The work contains a photo plots of arable dittany. Anatomical-morphological study of individual organs are shown in the photographs. The research was the structure of the secretory glands, Italian, ducts and conduits oils. Essential oil content was determined using three methods, and qualita- tive composition by GC/MS and SPME/GC/MS. Grown in the experimental plot plants reached an average height of 91 cm and 95 cm Albiflores variety Rosa Purple variety respectively. Flowering plants usually began in the third decade of May and continued until mid-June. The life of flowers is from 5 to 7 days. Flowers Albiflores varieties were characterized by a greater abundance of flowering and pollen higher efficiency and greater mass secreted nectar and honey. Analysis of the content and composition of the essential oil in the flowers cv. Albiflores content was 0.76%, while in the cv. Rosa Purple 0.60%, respectively 0.22% and 0.30% – herbs, and 0.10% and 0.02% roots. The main components of the essential oil obtained from the herb and roots were: limonene, myrcene, terpinolene, α-terpineol and germacrene D. KEY WORDS: DICTAMNUS ALBUS – ROSA PURPLE – ALBIFLORES – DITTANY – SECRETORY STRUCTURES – ESSENTIAL OIL
Transcript of Biologia wzrostu i rozwoju dwu odmian Dictamnus albus L ... · nego wykazały, że zachowanie tego...

21Postępy Fitoterapii 1/2015
Borgis Post Fitoter 2015; (16)1: 21-36
Prezentowana praca jest kontynuacją badań nad systematyką gatunku Dictamnus, jego rozmieszcze-niem w Europie i na świecie oraz przeglądem ważniej-szych grup związków biologicznie czynnych, ich bio-genezy, właściwości, występowania i zastosowania (1). W pierwszej części opisano alkaloidy furochinolinowe, furanokumaryny i limonoidy, zaś w drugiej omówiono olejki eteryczne, flawonoidy oraz inne związki biolo-gicznie aktywne (2, 3).
Dyptam jesionolistny znany jest pod nazwą „go-rejącego krzewu Mojżesza”. Nazwa ta pochodzi od opisu biblijnego rośliny: „Wtedy ukazał mu się Anioł Pański w płomieniu ognia, ze środka krzewu. Moj-żesz widział, jak krzew płonął ogniem, a nie spłonął od niego.” (Biblia, Księga Wyjścia). Dziś wiemy, że zjawisko to było możliwe gdyż wszystkie struktury w kwiatostanach dyptamu są gęsto pokryte gruczo-łami wydzielniczymi. Produkowane przez nie obficie olejki eteryczne w upalne, bezwietrzne dni mogą ulec samozapłonowi, nie uszkadzając rośliny (4, 5).
Postępujące skażenie środowiska naturalnego jest przyczyną zamierania wielu gatunków roślin, rosną-cych nawet na terenach parków narodowych i rezer-watów przyrody. Do takich ginących roślin należy zaliczyć Dictamnus albus L. (dyptam jesionolistny). Jako roślina reliktowa i bardzo rzadka, zawierająca ponadto szereg osobliwości strukturalnych i biologicz-nych, podlega ona całkowitej ochronie. Istnieje więc konieczność czynnej ochrony najbardziej zagrożonych populacji tych roślin w kontrolowanych warunkach. Mnożenie generatywne i uprawa dyptamu jesionolist-nego wykazały, że zachowanie tego gatunku w warun-kach ex situ daje pozytywne rezultaty. Dlatego też jest ona coraz częściej uprawiana jako roślina ozdobna o wyjątkowych walorach dekoracyjnych (6, 7).
*Tadeusz Wolski1,2, Tomasz Baj1
Biologia wzrostu i rozwoju dwu odmian dyptamu jesionolistnego (Dictamnus albus L. cv. Albiflores i cv. Rosa Purple) oraz budowa anatomiczno-morfologiczna struktur wydzielniczych, skład olejku eterycznego i analiza ekstraktu1Katedra i Zakład Farmakognozji z Pracownią Roślin Leczniczych, Uniwersytet Medyczny w Lublinie Kierownik Katedry i Zakładu: dr hab. Grażyna Zgórka 2Katedra Warzywnictwa i Roślin Leczniczych, Uniwersytet Przyrodniczy w Lublinie Kierownik Katedry: prof. dr hab. Jan Dyduch
BIOLOGY OF GROWTH AND DEVELOPMENT OF TWO CULTIVARS OF DICTAMNUS ALBUS L. (CV. ALBIFLORES AND CV. ROSA PURPLE), ANATOMICAL AND MORPHOLOGICAL STRUCTURE OF SECRETORY TISSUES AND ANALYSIS OF THE ESSENTIAL OIL
SUMMARYThe paper presents the biology of growth and development of two varieties of dittany (Dictamnus albus L. cv. Albiflores and cv. Rosa Purple). The present study include biometric measurements of morphological characteristics, including the flowering and pollination dynamics and efficiency of pollen. Measurements of the amount of shoots, stems of length and a width and length of the leaves. Set the number of flowering shoots, as well as the number of flowers on the shoot and the number of flowers per plant, given the dimensions of the grains of pollen, nectar and mass concentrations of sugar and honey yield. The work contains a photo plots of arable dittany. Anatomical-morphological study of individual organs are shown in the photographs. The research was the structure of the secretory glands, Italian, ducts and conduits oils. Essential oil content was determined using three methods, and qualita-tive composition by GC/MS and SPME/GC/MS. Grown in the experimental plot plants reached an average height of 91 cm and 95 cm Albiflores variety Rosa Purple variety respectively. Flowering plants usually began in the third decade of May and continued until mid-June. The life of flowers is from 5 to 7 days. Flowers Albiflores varieties were characterized by a greater abundance of flowering and pollen higher efficiency and greater mass secreted nectar and honey. Analysis of the content and composition of the essential oil in the flowers cv. Albiflores content was 0.76%, while in the cv. Rosa Purple 0.60%, respectively 0.22% and 0.30% – herbs, and 0.10% and 0.02% roots. The main components of the essential oil obtained from the herb and roots were: limonene, myrcene, terpinolene, α-terpineol and germacrene D.
KEY WORDS: DICTAMNUS ALBUS – ROSA PURPLE – ALBIFLORES – DITTANY – SECRETORY STRUCTURES – ESSENTIAL OIL

22
Tadeusz Wolski, Tomasz Baj
Postępy Fitoterapii 1/2015
Masę nektaru wydzieloną w okresie całego życia kwiatu określano metodą pipetową (12). Koncentra-cję cukrów w nektarze mierzono przy zastosowaniu refraktometru Abbego. Masę cukrów wytwarzaną w nektarze obliczono z 10, zaś wydajność miodową z 1000 kwiatów obu odmian (8, 9).
Badania anatomiczno-morfologiczne surowców
Badania anatomiczno-morfologiczne obejmowały nadziemne i podziemne części dwu odmian dyptamu jesionolistnego. (13-16).
Tkanki badano na przekrojach poprzecznych wyko-nanych ze świeżego oraz utrwalonego w FFA (etanol 50% + kwas octowy lodowaty + formalina 40%; 90:5:5) materiału roślinnego (17). Zastosowano testy histochemiczne w celu zabarwienia struktur komórko-wych i eksudatów tkanki wydzielniczej w skrawkach uzyskanych ze świeżych organów: płyn Lugola, Sudan III oraz błękit toluidynowy.
Fragmenty kwiatostanów, liści, łodyg i korzeni utrwalano w 2,5% aldehydzie glutarowym w 0,1 M buforze kokodylanowym (pH = 7,4). Po dehydratacji próbki przenoszono do tlenku propylenu i zatapiano w żywicy SPURR. Skrawki o grubości 0,5-0,7 µm uzyskiwano przy użyciu ultramikrotomu Reichert Ultracut S.
Po utrwaleniu w aldehydzie glutarowym i odwod-nieniu, część próbek przenoszono do acetonu. Na-stępnie suszono je w ciekłym CO2 i napylano złotem przy użyciu CS 100 Sputter Coater. Obserwacje wy-konywano przy użyciu mikroskopu BS-340 Tesla. Do barwienia stosowano 1% roztwory: błękitu metyleno-wego z azurem II w wodnym roztworze czteroboranu sodu.
Olejek eterycznyProcentową zawartość olejku eterycznego określano
zgodnie z FP IV (18), FP V, (19, 20) i PN 91/R-87019 (21). Olejki eteryczne oznaczano metodą destylacji z parą wodną w aparacie Derynga trzema metodami: bezpośrednio – metoda I, pośrednio z m-ksylenem – metoda II, oraz po hydrolizie badanych surowców przy użyciu 20% kwasu siarkowego – metoda III.
Wykonanie oznaczeń
Metoda I – destylacja bezpośrednia z parą wodną: próbki (w trzech powtórzeniach) odpowiednio roz-drobnionych surowców (liście i kwiatostany – 20 g, rozkruszone; kłącza – 10 g, sito 0,28 mm) umiesz-czano w kolbach o pojemności 1000 ml i dodawano wodę destylowaną w proporcji 1/20 (surowiec/woda), następnie po połączeniu kolby z aparatem Derynga i napełnieniu odbieralnika wodą, włączano chłodze-
W piśmiennictwie florystycznym i ogrodniczym dotyczącym dyptamu jesionolistnego podawana jest charakterystyka głównie części nadziemnych, a zwłaszcza liści i kwiatów, z pominięciem organów podziemnych i rytmiki rozwojowej tej rośliny. Celem prezentowanej pracy były badania nad biologią wzro-stu i rozwoju dyptamu jesionolistnego. Przedmiotem badań prezentowanej pracy były organy nadziemne i podziemne (kwiaty, ziele, torebki nasienne, nasiona, korzeń). Badania obejmowały ocenę biologii wzrostu i rozwoju dyptamu jesionolistnego, w tym pomiary biometryczne, dynamikę kwitnienia i pręcikowania oraz charakterystykę budowy morfologiczno-anato-micznej, ze szczególnym uwzględnieniem struktur wydzielniczych, oraz zawartość i skład olejku ete-rycznego.
Pomiary biometryczne cech morfologicznych dyptamu jesionolistnego
Wszystkie pomiary biometryczne prowadzono na 10 losowo wybranych okazach roślin, każdej z od-mian dyptamu jesionolistnego; pomiary powtarzano 6-krotnie, w tygodniowych odstępach czasu. Pomiary obejmowały: wysokość pędów głównych (każdorazo-wo po 20 dla każdej odmiany), długość łodyg bocz-nych (każdorazowo po 20 dla każdej odmiany), liczbę listków na łodygach bocznych (każdorazowo po 30 dla każdej odmiany), długość i szerokość listków na łodygach bocznych (każdorazowo po 30 dla każdej odmiany).
Dzienna dynamika rozkwitania i pyłkowania
Dzienną dynamikę rozkwitania określano dla 18 kwiatostanów każdej odmiany (3 kwiatostany w 6 roślinach). Liczbę rozkwitłych kwiatów obserwowano co godzinę od 6:30 do 17:30. Wyniki przedstawiono po przeliczeniu na udział procentowy kwiatów roz-kwitłych w ciągu godziny w stosunku do wszystkich kwiatów, które rozwinęły się w ciągu dnia (100%). Porównywano długość życia kwiatów obu odmian. Pod koniec okresu kwitnienia wykonywano pomiary długości 70 pędów kwiatostanowych, a liczbę kwiatów na pędzie ustalano dla 60 pędów każdej rośliny. Do określenia liczby kwitnących pędów na roślinie wyko-rzystywano po 13 roślin obu odmian (8, 9).
Wydajność pyłkową badano oddzielnie dla kwia-tów położonych na osi głównej pędu oraz na od-gałęzieniach drugiego rzędu. Zastosowano metodę eterowo-wagową Warakomskiej (10) z modyfikacją Szklanowskiej i Pluty (11). Wymiary 200 ziaren pyłku dla każdej odmiany badano w preparatach gliceroże-latynowych.
23
Biologia wzrostu i rozwoju dwu odmian dyptamu jesionolistnego (Dictamnus albus L. cv. Albiflores i cv. Rosa Purple)...
Postępy Fitoterapii 1/2015
D. albus. Surowce bezpośrednio po zbiorze umieszcza-no w próbówce, a następnie lotne składniki kwiatosta-nu poddawano sorpcji na włóknie strzykawki w czasie 30 min. Zaadsorbowane składniki olejku eterycznego analizowano metodą GC/MS.
Analiza GC/MS olejku eterycznego
Analizę GC/MS prowadzono przy użyciu aparatu ITS-40 (układ GC/ITMS firmy Finnigan MAT, USA) z kolumną DB-5 (J&W, USA) o długości 30 m, średni-cy 0,25 mm i grubości filmu fazy stacjonarnej 0,25 µm. Temperatura dozownika 280°C. Stosowano gradient temperatury (35°C przez 2 min, następnie przyrost co 4°C do 280°C). Analizę jakościową przeprowadzano na podstawie widm MS, porównując je z widmami bi-blioteki NIST (62 tys. widm), oraz biblioteki terpenów LIBR (TR) ( Finnigan MAT). Tożsamość związków potwierdzano na podstawie indeksów zamieszczonych w piśmiennictwie (22) i własnych.
Wyniki i omówienie
Pomiary biometryczne cech morfologicznych
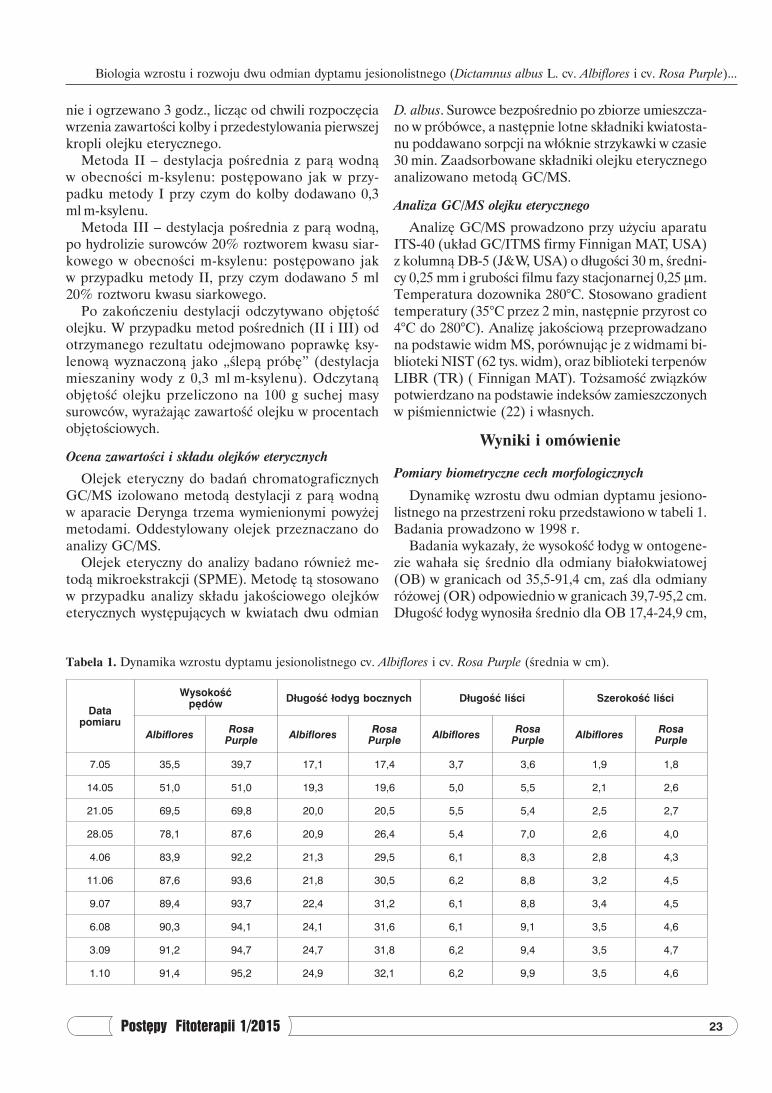
Dynamikę wzrostu dwu odmian dyptamu jesiono-listnego na przestrzeni roku przedstawiono w tabeli 1. Badania prowadzono w 1998 r.
Badania wykazały, że wysokość łodyg w ontogene-zie wahała się średnio dla odmiany białokwiatowej (OB) w granicach od 35,5-91,4 cm, zaś dla odmiany różowej (OR) odpowiednio w granicach 39,7-95,2 cm. Długość łodyg wynosiła średnio dla OB 17,4-24,9 cm,
nie i ogrzewano 3 godz., licząc od chwili rozpoczęcia wrzenia zawartości kolby i przedestylowania pierwszej kropli olejku eterycznego.
Metoda II – destylacja pośrednia z parą wodną w obecności m-ksylenu: postępowano jak w przy-padku metody I przy czym do kolby dodawano 0,3 ml m-ksylenu.
Metoda III – destylacja pośrednia z parą wodną, po hydrolizie surowców 20% roztworem kwasu siar-kowego w obecności m-ksylenu: postępowano jak w przypadku metody II, przy czym dodawano 5 ml
20% roztworu kwasu siarkowego.Po zakończeniu destylacji odczytywano objętość
olejku. W przypadku metod pośrednich (II i III) od otrzymanego rezultatu odejmowano poprawkę ksy-lenową wyznaczoną jako „ślepą próbę” (destylacja mieszaniny wody z 0,3 ml m-ksylenu). Odczytaną objętość olejku przeliczono na 100 g suchej masy surowców, wyrażając zawartość olejku w procentach objętościowych.
Ocena zawartości i składu olejków eterycznych
Olejek eteryczny do badań chromatograficznych GC/MS izolowano metodą destylacji z parą wodną w aparacie Derynga trzema wymienionymi powyżej metodami. Oddestylowany olejek przeznaczano do analizy GC/MS.
Olejek eteryczny do analizy badano również me-todą mikroekstrakcji (SPME). Metodę tą stosowano w przypadku analizy składu jakościowego olejków eterycznych występujących w kwiatach dwu odmian
Tabela 1. Dynamika wzrostu dyptamu jesionolistnego cv. Albiflores i cv. Rosa Purple (średnia w cm).
Data pomiaru
Wysokośćpędów Długość łodyg bocznych Długość liści Szerokość liści
Albiflores Rosa Purple Albiflores Rosa
Purple Albiflores Rosa Purple Albiflores Rosa
Purple
7.05 35,5 39,7 17,1 17,4 3,7 3,6 1,9 1,8
14.05 51,0 51,0 19,3 19,6 5,0 5,5 2,1 2,6
21.05 69,5 69,8 20,0 20,5 5,5 5,4 2,5 2,7
28.05 78,1 87,6 20,9 26,4 5,4 7,0 2,6 4,0
4.06 83,9 92,2 21,3 29,5 6,1 8,3 2,8 4,3
11.06 87,6 93,6 21,8 30,5 6,2 8,8 3,2 4,5
9.07 89,4 93,7 22,4 31,2 6,1 8,8 3,4 4,5
6.08 90,3 94,1 24,1 31,6 6,1 9,1 3,5 4,6
3.09 91,2 94,7 24,7 31,8 6,2 9,4 3,5 4,7
1.10 91,4 95,2 24,9 32,1 6,2 9,9 3,5 4,6
24
Tadeusz Wolski, Tomasz Baj
Postępy Fitoterapii 1/2015
Liczba rozkwitających KwR osiągnęła dwa maksima – pomiędzy 10:30 a 12:30 i 14:30 a 17:30. W godzinach popołudniowych następowało zahamowanie otwie-rania się kwiatów. W dniu prowadzenia obserwacji notowano stopniowy wzrost dziennej temp. od 14 do 25°C. W przypadku KwB także zaobserwowano dwa okresy maksymalnego rozkwitania w ciągu dnia: pierwszy 7:30-9:30, drugi 13:30-16:30. W przypadku obu odmian w południe następował spadek liczby rozwijających się kwiatów (8).
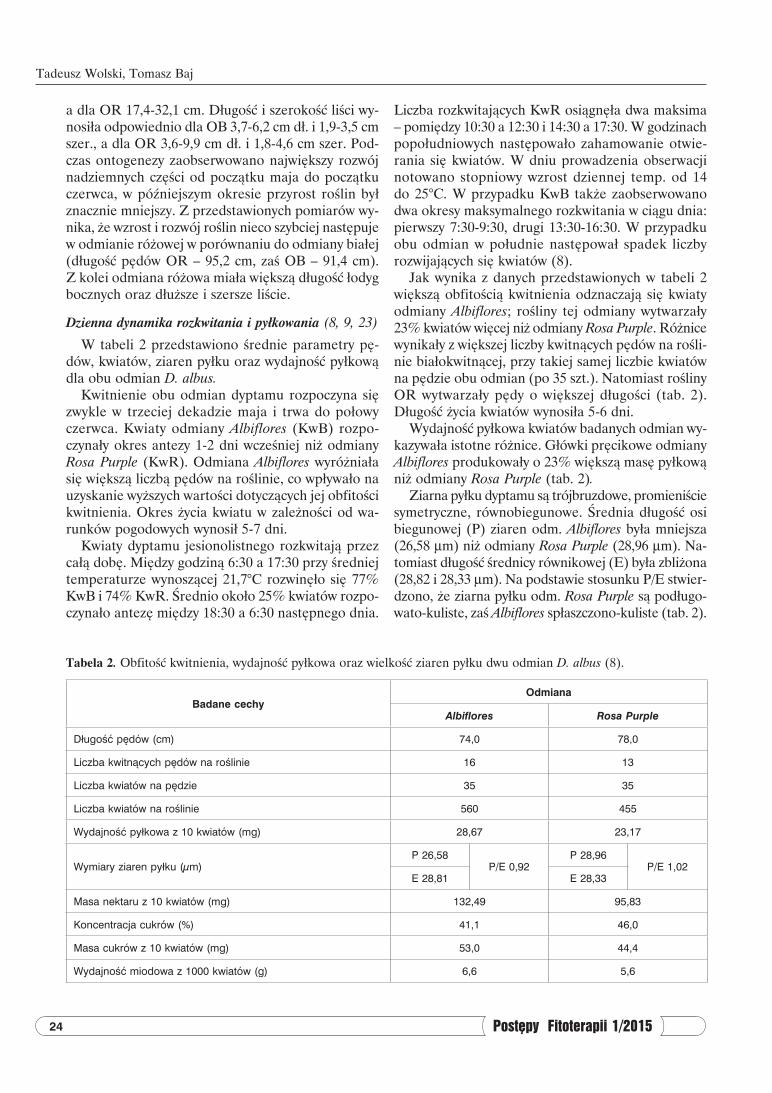
Jak wynika z danych przedstawionych w tabeli 2 większą obfitością kwitnienia odznaczają się kwiaty odmiany Albiflores; rośliny tej odmiany wytwarzały 23% kwiatów więcej niż odmiany Rosa Purple. Różnice wynikały z większej liczby kwitnących pędów na rośli-nie białokwitnącej, przy takiej samej liczbie kwiatów na pędzie obu odmian (po 35 szt.). Natomiast rośliny OR wytwarzały pędy o większej długości (tab. 2). Długość życia kwiatów wynosiła 5-6 dni.
Wydajność pyłkowa kwiatów badanych odmian wy-kazywała istotne różnice. Główki pręcikowe odmiany Albiflores produkowały o 23% większą masę pyłkową niż odmiany Rosa Purple (tab. 2).
Ziarna pyłku dyptamu są trójbruzdowe, promieniście symetryczne, równobiegunowe. Średnia długość osi biegunowej (P) ziaren odm. Albiflores była mniejsza (26,58 µm) niż odmiany Rosa Purple (28,96 µm). Na-tomiast długość średnicy równikowej (E) była zbliżona (28,82 i 28,33 µm). Na podstawie stosunku P/E stwier-dzono, że ziarna pyłku odm. Rosa Purple są podługo-wato-kuliste, zaś Albiflores spłaszczono-kuliste (tab. 2).
a dla OR 17,4-32,1 cm. Długość i szerokość liści wy-nosiła odpowiednio dla OB 3,7-6,2 cm dł. i 1,9-3,5 cm szer., a dla OR 3,6-9,9 cm dł. i 1,8-4,6 cm szer. Pod-czas ontogenezy zaobserwowano największy rozwój nadziemnych części od początku maja do początku czerwca, w późniejszym okresie przyrost roślin był znacznie mniejszy. Z przedstawionych pomiarów wy-nika, że wzrost i rozwój roślin nieco szybciej następuje w odmianie różowej w porównaniu do odmiany białej (długość pędów OR – 95,2 cm, zaś OB – 91,4 cm). Z kolei odmiana różowa miała większą długość łodyg bocznych oraz dłuższe i szersze liście.
Dzienna dynamika rozkwitania i pyłkowania (8, 9, 23)
W tabeli 2 przedstawiono średnie parametry pę-dów, kwiatów, ziaren pyłku oraz wydajność pyłkową dla obu odmian D. albus.
Kwitnienie obu odmian dyptamu rozpoczyna się zwykle w trzeciej dekadzie maja i trwa do połowy czerwca. Kwiaty odmiany Albiflores (KwB) rozpo-czynały okres antezy 1-2 dni wcześniej niż odmiany Rosa Purple (KwR). Odmiana Albiflores wyróżniała się większą liczbą pędów na roślinie, co wpływało na uzyskanie wyższych wartości dotyczących jej obfitości kwitnienia. Okres życia kwiatu w zależności od wa-runków pogodowych wynosił 5-7 dni.
Kwiaty dyptamu jesionolistnego rozkwitają przez całą dobę. Między godziną 6:30 a 17:30 przy średniej temperaturze wynoszącej 21,7°C rozwinęło się 77% KwB i 74% KwR. Średnio około 25% kwiatów rozpo-czynało antezę między 18:30 a 6:30 następnego dnia.
Tabela 2. Obfitość kwitnienia, wydajność pyłkowa oraz wielkość ziaren pyłku dwu odmian D. albus (8).
Badane cechyOdmiana
Albiflores Rosa Purple
Długość pędów (cm) 74,0 78,0
Liczba kwitnących pędów na roślinie 16 13
Liczba kwiatów na pędzie 35 35
Liczba kwiatów na roślinie 560 455
Wydajność pyłkowa z 10 kwiatów (mg) 28,67 23,17
Wymiary ziaren pyłku (µm)P 26,58
P/E 0,92P 28,96
P/E 1,02E 28,81 E 28,33
Masa nektaru z 10 kwiatów (mg) 132,49 95,83
Koncentracja cukrów (%) 41,1 46,0
Masa cukrów z 10 kwiatów (mg) 53,0 44,4
Wydajność miodowa z 1000 kwiatów (g) 6,6 5,6

25
Biologia wzrostu i rozwoju dwu odmian dyptamu jesionolistnego (Dictamnus albus L. cv. Albiflores i cv. Rosa Purple)...
Postępy Fitoterapii 1/2015
czenie gruczołów jest u obydwu odmian podobne. Górna powierzchnia płatków korony i szyjka słupka odznaczają się ich brakiem. Największe skupienie gruczołów u odmiany białej występuje na łodyżkach, liściach przykwiatowych i działkach kielicha (dolna powierzchnia), na zalążni słupka oraz górnej części nitek pręcikowych, podobnie jak u odmiany różowej (ryc. 1 i 2).
Wielkość gruczołów wydzielniczych u porównywa-nych odmian jest zbliżona. Dla odmiany białej więk-szość z nich składa się z wielokomórkowego trzonu i najczęściej owalnej główki, której centralną część wypełniają komórki wydzielnicze lub tworzący się z czasem zbiornik wydzielniczy. Obserwowano gru-czoły w różnych stadiach rozwojowych. W młodszych gruczołach wewnętrzną część stanowiły luźno ułożo-ne komórki wydzielnicze, które stopniowo ulegały rozpadowi. W starszych gruczołach, komórki epitelu otaczały zbiornik olejkowy.
W położeniu biegunowym ziarna są kulisto-trójkąt-ne, a w równikowym lekko owalne. W obrębie bruzd położone są kuliste lub wydłużone równikowo pory. Powierzchnia egzyny odznacza się nieregularnie roz-mieszczonymi prążkami.
W kwiatach D. albus występuje 10 pręcików. W cza-sie antezy nitki pręcikowe zmieniają swoje położenie, kształt i wymiary. W górnej części nitek występują gru-czoły wydzielnicze wytwarzające intensywny zapach. W pąku pręciki tworzą zwartą wiązkę wokoło słupka, położoną poziomo lub lekko odchyloną ku dołowi. Równocześnie z rozchylaniem się płatków korony, nit-ki pręcików górnej części wiązki stopniowo wydłużają się, lekko podnoszą, a następnie wyginają łukowato unosząc ku górze główki. W tej pozycji pylniki pękają bocznie, uwalniając ziarna pyłku. Proces dojrzewania jednego pręcika obejmuje elongację i wygięcie nitki oraz początek pylenia.
Masa nektaru wydzielonego z 10 kwiatów w fazie pełni kwitnienia roślin kształtowała się średnio na poziomie 132,49 mg dla KwB, natomiast dla odmiany KwR wynosiła 95,83. Zaobserwowano wyższe stęże-nie cukrów w nektarze KwR, przy czym ich masa 10 kwiatów była wyższa dla KwB. Wydajność miodowa 1000 kwiatów odmiany Albiflores była nieco wyższa i wynosiła 6,6 g, zaś dla odmiany Rosa Purple wynosiła ona 5,6 g.
Badania anatomiczno-morfologiczne surowców
W piśmiennictwie brak jest systematycznych ba-dań anatomiczno-morfologicznych. W Polsce ostanie obserwacje nad budową anatomiczno-morfologiczną prowadzone były przez Gertiga i wsp. (24) i dotyczyły one roślin występujących w stanie naturalnym. Pre-zentowane obserwacje obejmowały ocenę budowy morfologiczno-anatomicznej dwu odmian dyptamu jesionolistnego uprawianego na poletku doświad-czalnym Katedry Roślin Ozdobnych AR w Lublinie (ryc. 1), ze szczególnym uwzględnieniem lokalizacji struktur wydzielniczych. Badania były wykonane we współpracy z Katedrą Botaniki Wydziału Ogrodni-czego AR w Lublinie.
Kwiaty dyptamu odm. Albiflores i odm. Rosa Purple różnią się nie tylko barwą płatków korony, lecz także zabarwieniem szypułek, liści przykwiatowych oraz gruczołów wydzielniczych występujących w obrębie kwiatostanów. U odmiany o kwiatach białych szy-pułki i liście towarzyszące kwiatom są zielone, a gru-czoły wydzielnicze zielonożółte, natomiast odmiana o kwiatach różowych ma wymienione części kwiato-stanów w kolorze brunatnobordowym, zaś gruczoły wydzielnicze mają zwykle barwę ciemnoczerwoną lub znacznie rzadziej barwę czerwonozieloną. Rozmiesz-
Ryc. 1. Szczytowa część kwiatostanu dyptamu z silnie ogróczo-lonymi łodyżkami i pąkami kwiatowymi cv. Rosa Purple (1).
Ryc. 2. Kwiaty dyptamu cv. Albiflores z widocznymi gruczołami wydzielniczymi (pow. 1,5x) (34).

26
Tadeusz Wolski, Tomasz Baj
Postępy Fitoterapii 1/2015
wydzielnicze, występujące na pozostałych elementach kwiatostanu, odznaczają się różnorodnością struktury. Najczęściej składają się one z wielorzędowego trzonu i owalnej wielokomórkowej główki, zaopatrzonej na szczycie w delikatny kilkukomórkowy włosek. Wło-sek ten łatwo ulega uszkodzeniu, czemu towarzyszy uwalnianie wydzieliny. Gruczoły ekskrecyjne różnią się przede wszystkim wielkością główki i długością trzonu.
Nasiona są koloru czarnego, mają kształt gruszko-waty. Są one połyskujące i gładkie. Ich długość wynosi średnio 5 mm, zaś grubość w najszerszym miejscu 2,5 mm. Nasiona mają białe znamię na zwężonym końcu. Okrywa nasienna jest dość gruba, krucha, pod nią znajduje się białe bielmo. Zarodek jest drob-ny, ma dwa cienkie liścienie i walcowaty korzonek (ryc. 5-7).
Liście właściwe osadzone są na łodydze skręto-legle. Są one ciemnozielone, złożone, nieparzysto-pierzaste, najczęściej 7-9-listkowe. Pojedyncze listki
Na szczycie główki gruczołu w odmianie różowej i białej występuje delikatny, ostro zakończony, kil-kukomórkowy włosek. Przenikanie olejków eterycz-nych na zewnątrz odbywa się prawdopodobnie przez cienkie ściany komórek epidermy w główce gruczołu. Między komórkami epidermy na trzonach włosków obserwowano zagłębienia z niezbyt intensywnie zabar-wioną zawartością, które być może stanowią również miejsca uwalniania wydzieliny.
Zewnętrzne gruczoły wydzielnicze występują w szczególnie dużym zwarciu na łodydze kwiatosta-nowej, szypułkach i zalążni słupka oraz odosiowej powierzchni liści przykwiatowych (ryc. 3 i 4).
Obliczono, że na osi kwiatostanu występuje około 9 gruczołów na 1 mm2 powierzchni epidermy. Gru-czoły położone w górnej części nitek pręcikowych charakteryzują się zbliżonym kształtem i wielkością. Ich wysokość wynosi około 330 µm, natomiast gruczoły
Ryc. 3. Gruczoły wydzielnicze na powierzchni zalążni słupka dyptamu (pow. 200x).
Ryc. 4. Gruczoł wydzielniczy z widocznym włoskiem na szczycie z powierzchni płatka korony (pow. 300x).
Ryc. 5. Dojrzała pięciokomorowa torebka dyptamu (pow. 8x).
Ryc. 6. Fragment przekroju poprzecznego dojrzewającej torebki dyptamu z widocznymi nasionami oraz gruczołami wydzielniczymi (pow. 24x).

27
Biologia wzrostu i rozwoju dwu odmian dyptamu jesionolistnego (Dictamnus albus L. cv. Albiflores i cv. Rosa Purple)...
Postępy Fitoterapii 1/2015
Zbiorniki wydzielnicze zawierające kilka warstw komórek, tworzą epitel gruczołowy i centralnie po-łożoną przestrzeń, w której gromadzi się olejek. Są one większe niż w łodydze, a ich średnica wynosi około 100 µm.
Grupa komórek epidermy (w liczbie 7-10) poło-żonych nad zbiornikiem wydzielniczym odznacza się zwartym układem. Ich ściany okrywa gładka warstwa kutykuli, w odróżnieniu od pozostałych komórek epidermy o kutykuli prążkowanej. W skórce górnej nie stwierdzono obecności aparatów szparkowych. W epidermie odosiowej liści obserwowano liczne aparaty szparkowe usytuowane na niewielkich wynie-sieniach epidermy. Obliczono, że na 1 mm2 epidermy liści dyptamu występuje 125 szparek.
Zbiorniki olejkowe w blaszkach liściowych dyptamu położone są tylko przy powierzchni doosiowej. Wystę-pują one także w nerwach i ogonkach liściowych oraz w łodygach. Składają się one z centralnie położonej komory oraz otaczającej ją warstwy wydzielniczej epitelialnej, obejmującej 1-3 pokłady komórek. Mię-kisz palisadowy formuje jedną warstwę zajmującą około 1/3 grubości blaszki. Miękisz gąbczasty jest 7-8-warstwowy i wykazuje obecność bardzo dużych przestworów międzykomórkowych. Odosiowa epider-ma wyposażona jest w aparaty szparkowe, tworzące niewielkie wyniesienia powierzchni tej tkanki.
Zbiorniki wydzielnicze rozmieszczone są w mięki-szu palisadowym. Ich budowa jest taka sama jak zbior-ników występujących w łodygach i nerwach liści. Ze względu na sposób powstawania można je zaliczyć do schizolizygenicznych. Komórki epidermy nad zbiorni-kami są bardziej spłaszczone i mniejsze niż pozostałe, co sprawia, że w powierzchni liścia powstają niewielkie zagłębienia. Zewnętrzne ściany komórkowe epidermy w tej strefie są znacznie cieńsze niż pozostałych ko-mórek skórki, co niewątpliwie pozostaje w związku z procesem odparowywania olejków eterycznych.
Na rycinach 8-14 przedstawione zostały zbiorniki wydzielnicze na przekrojach nerwu głównego i liści oraz na powierzchni liści dyptamu.
W obrębie kwiatostanu łodyga pokryta jest liczny-mi gruczołami wydzielniczymi. W niżej położonych częściach rośliny jej powierzchnia pozbawiona jest zewnętrznych gruczołów wydzielniczych. Łodyga ma kształt obły lub zbliżony do trójkątnego. W obłej części łodygi wyraźnie zaznaczają się dwa żeberka.
Łodyga w górnej części kwiatostanu na przekroju poprzecznym wykazuje budowę pierwotną z wyspo-wym rozmieszczeniem wiązek przewodzących, oddzie-lonych promieniami rdzeniowymi. W pozostałych czę-ściach łodygi obserwuje się budowę wtórną. Komórki epidermy na przekroju poprzecznym są czworokątne
o owalnym kształcie i brzegu piłkowanym położone są na oskrzydlonej osadce przechodzącej w oskrzy-dlony ogonek.
Blaszki liściowe oglądane makroskopowo nie ujaw-niają obecności zbiorników olejkowych, które są małe i występują nielicznie w mezofilu liścia. Położenie zbiorników można dostrzec oglądając powierzchnię utrwalonej blaszki liściowej na małych powiększeniach mikroskopowych. Stwierdzono, że zbiorniki te są regularnie rozmieszczone przy brzegu blaszki, obok nasady piłkowanych występów. W innych miejscach zbiorniki olejkowe są bardziej oddalone od siebie i występują w liczbie 1-3 na 1 mm2 obserwowanej powierzchni liścia.
Nerw główny liści właściwych tworzy masywne uwypuklenie po stronie odosiowej. Okrywająca go epiderma składa się z drobnych, owalnych komórek o zewnętrznych ścianach zgrubiałych, z warstwą ku-tykuli.
W środkowej strefie zgrubienia nerwu położone są różnej wielkości wiązki przewodzące (w liczbie 1-4). Górną i dolną część nerwu wzmacnia kolenchyma kątowa tworząca kilka warstw.
W obrazie mikroskopowym blaszek liściowych widoczne są dość nierównomiernie rozmieszczone zbiorniki olejkowe o różnej średnicy. Z obliczeń wy-nika, że liczba zbiorników przypadająca na 1 mm2 powierzchni liścia dyptamu wynosi 1-5, średnio 2,5. Długość średnicy zbiorników ekskrecyjnych zawarta była w przedziale 52-93 µm, średnio 72 µm. Obserwa-cje mikroskopowe ujawniły występowanie zagłębień w blaszce liściowej, które towarzyszą zbiornikom wydzielniczym. Zagłębienia te widoczne są również w brzeżnej części blaszki, u nasady piłkowanych wcięć.
Ryc. 7. Gruczoły wydzielnicze na powierzchni dojrzewającego owocu (pow. 25x).

28
Tadeusz Wolski, Tomasz Baj
Postępy Fitoterapii 1/2015
Ryc. 8. Przekrój poprzeczny nerwu głównego liścia dyptamu. W dolnej części widoczny zbiornik wydzielniczy (strzałka) (pow. 45x).
Ryc. 9. Fragment przekroju poprzecznego nerwu głównego liścia dyptamu ze zbiornikiem wydzielniczym (strzałka) (pow. 110x).
Ryc. 10. Fragment brzeżnej części blaszki liściowej dyptamu z widocznym zagłębieniem nad zbiornikiem wydzielniczym (strzałka) (pow. 170x) (14).
Ryc. 12. Przekrój poprzeczny liścia dyptamu z widocznym zbiornikiem wydzielniczym (pow. 220x).
Ryc. 11. Fragment dolnej powierzchni liścia dyptamu z widocz-nymi aparatami szparkowymi (pow. 340x).
Ryc. 13. Przekrój poprzeczny blaszki liściowej dyptamu. Wi-doczny zbiornik wydzielniczy między komórkami miękiszu palisadowego (strzałka) (pow. 220x) (15).

29
Biologia wzrostu i rozwoju dwu odmian dyptamu jesionolistnego (Dictamnus albus L. cv. Albiflores i cv. Rosa Purple)...
Postępy Fitoterapii 1/2015
wytwarzając pustą część środkową, w której gromadzi się wydzielina. Miękisz zieleniowy łodygi składa się z 3-5 warstw komórek niewielkich rozmiarów, które sąsiadują z miękiszem bezzieleniowym o komórkach kilkakrotnie większych. Endoderma stanowi poje-dynczą warstwę komórek z licznymi amyloplastami, zawierającymi ziarna skrobi. Na rycinie 16 przed-stawiono gruczoły wydzielnicze zlokalizowane na szypułce kwiatowej.
Łyko wtórne w zewnętrznej części wytwarza pasma włókien sklerenchymatycznych. Drewno wtórne for-muje przyrost jednolity z promieniami rdzeniowymi 1-3-rzędowymi. W skład ksylemu wchodzą naczynia, cewki, miękisz i włókna drzewne. Centralną część łodygi wypełnia rozległy rdzeń miękiszowy.
Część podziemną D. albus stanowi dość rozbudo-wany system korzeniowy. Z hypokotylu wyrastają dość licznie korzenie przybyszowe o średniej długości 50 cm i średnicy 4 cm. (ryc. 17). Na grubszych korzeniach zaobserwowano liczne drobne korzonki boczne, które tworzyły gęste sploty. Powierzchnia korzeni jest mato-wa o barwie jasnożółtej. Na przekrojach poprzecznych wyróżnić możemy: skórkę, szeroki pas kory pierwot-nej oraz walec osiowy. Komórki skórki są kształtu nieregularnego o różnej wielkości. Kora pierwotna
lub owalne. Zewnętrzna część ściany komórkowej jest lekko uwypuklona na zewnątrz i zgrubiała, pokryta warstwą kutykuli o grubości zbliżonej do zgrubień ściany. Aparaty szparkowe są wyniesione powyżej po-ziomu epidermy przez duże komórki przyszparkowe. Kolenchyma występująca pod epidermą tworzy od 1 do 4 warstw. Nie stanowi ona warstwy ciągłej, jest przedzielona miękiszem.
Niewielkie zbiorniki wydzielnicze o średnicy 60-80 µm położone są w uwypuklonych częściach ło-dyg, bezpośrednio pod epidermą (ryc. 15). Komórki epidermy położone nad zbiornikami są mniejsze i bardziej spłaszczone niż inne komórki tej tkanki. W centralnej części zbiorników często obserwowano słabo zarysowujące się kontury komórek sekrecyj-nych, które prawdopodobnie stopniowo ulegają lizie
Ryc. 14. Zbiornik wydzielniczy (z) na przekroju poprzecznym nerwu głównego liścia dyptamu (pow. 420x) (15).
Ryc. 15. Fragmenty przekrojów poprzecznych łodygi dyptamu z widocznymi zbiornikami wydzielniczymi (strzałka) (A – pow. 110x; B – pow. 200x) (5).
A B
Ryc. 16. Gruczoły wydzielnicze na przekroju poprzecznym szypułki kwiatowej (pow. 220x).
Ryc. 17. System korzeniowy dyptamu jesionolistnego cv. Albi-flores; wygląd makroskopowy.

30
Tadeusz Wolski, Tomasz Baj
Postępy Fitoterapii 1/2015
między odmianami stwierdzić można, że odmiana białokwiatowa zawiera go nieco więcej niż odmiana różowo kwiatowa.
W tabeli 4 przedstawiono zawartość procentową olejku eterycznego w owocostanach (torebkach na-siennych) oznaczoną trzema metodami.
Przedstawione badania wskazują, że hydroliza kwaśna uwolniła część składników olejku eteryczne-go związanych glikozydowo. Zawartość procentowa olejku w owocostanach obu badanych odmian była najwyższa po destylacji surowca metodą III.
zbudowana jest z komórek owalnych. Ułożone są one regularnie, najmniejsze z nich znajdują się w zewnętrz-nych warstwach kory pierwotnej. Naczynia pierwotne ułożone są promieniście w podwójne szeregi oddzie-lone od siebie pojedynczą, podwójną, bądź potrójną warstwą komórek miękiszowych. W walcu osiowym wyróżnić możemy dość duże promienie rdzeniowe. Na przekrojach poprzecznych o budowie wtórnej wyróżnić można: perydermę, pas kory pierwotnej, łyko oraz drewno. W komórkach miękiszu kory pierwotnej występują dość licznie ziarna skrobi i komórki gruzłów szczawianu wapnia (ryc. 18, 19).
Analiza olejku eterycznegoZawartość procentową olejku eterycznego (OE)
w kwiatach (Kw), zielu (Z) i korzeniach (Ko) dwu odmian dyptamu jesionolistnego: biało kwiatowej (B) i różowo kwiatowej (R) przedstawiono w tabeli 3.
Jak wynika z tabeli 3 najwyższą zawartość pro-centową olejku eterycznego stwierdzono w kwia-tach; średnia zawartość w KwB wynosiła 0,76%, zaś w KwR 0,60%. W zielu i korzeniach obu odmian średnia procentowa zawartość olejku była na zbliżo-nym poziomie: ZB 0,22%, KoB 0,10%, ZR 0,20% i KoR 0,12%. Z porównania zawartości olejku po-
Ryc. 18, 19. Przekroje poprzeczne przez korzenie dyptamu jesionolistnego, A – ziarna skrobi, B – gruzły szczawianu wapnia, C – promienie rdzeniowe (ryc. 18 – pow. 110x; ryc. 19 – pow. 200x) (26, 27).
Tabela 3. Procentowa zawartość olejku eterycznego w badanych surowcach w przeliczeniu na suchą masę (34).
Data zbioruZawartość olejku eterycznego (%)
KwB ZB KoB KwR ZR KoR
5.06.1999 0,73 0,19 0,09 0,60 0,18 0,10
10.06.2000 0,75 0,23 0,10 0,58 0,20 0,11
1.06.2001 0,80 0,24 0,12 0,63 0,22 0,14
Średnia 0,76 ± 0,04 0,22 ± 0,03 0,1 ± 0,02 0,6 ± 0,03 0,2 ± 0,02 0,17 ± 0,02
Tabela 4. Zawartość procentowa olejków eterycznych w owocostanach oznaczona trzema metodami (35).
Metoda destylacjiBadana odmiana
cv. Albiflores cv. Rosa Purple
I 0,12 0,08
II 0,12 0,09
III 0,16 0,12
Metody destylacji bezpośredniej z parą wodną: I – bezpośrednia, II – z m-ksylenem, III – z m-ksylenem i 20% kwasem siarkowym

31
Biologia wzrostu i rozwoju dwu odmian dyptamu jesionolistnego (Dictamnus albus L. cv. Albiflores i cv. Rosa Purple)...
Postępy Fitoterapii 1/2015
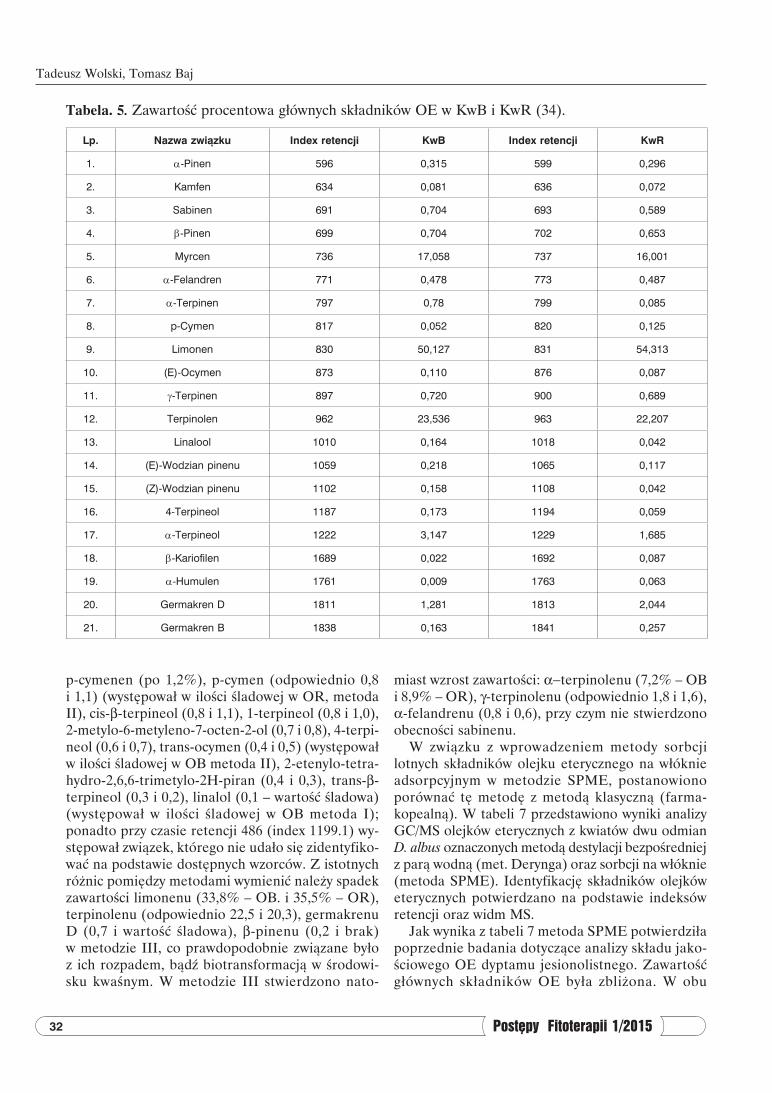
nen, terpinolen oraz myrcen. Analizując skład olejku eterycznego KwB i KwR zidentyfikowano ogółem 21 związków chemicznych. W odmianie białokwiatowej w porównaniu do odmiany różowej stwierdzono nieco większą zawartość terpinolenu (odpowiednio 23,54 wobec 22,21%) i myrcenu (17,06% wobec 16,00%), na-tomiast zawartość limonenu w KwB wynosiła 50,13%, zaś w KwR 54,31%. Wśród składników występujących powyżej 0,5% zidentyfikowano w obu odmianach: α-terpineol (KwB – 3,15%; KwR – 1,69%), germakren D (odpowiednio 1,28 i 2,04%), γ-terpinolen (odpowied-nio 0,72 i 0,69%), β-pinen (odpowiednio 0,70 i 0,65%), sabinen (odpowiednio 0,70 i 0,59%). Ponadto odmiana KwB zawierała α-terpinen w ilości 0,78%, podczas gdy KwR zawierała 0,085% tego związku (35).
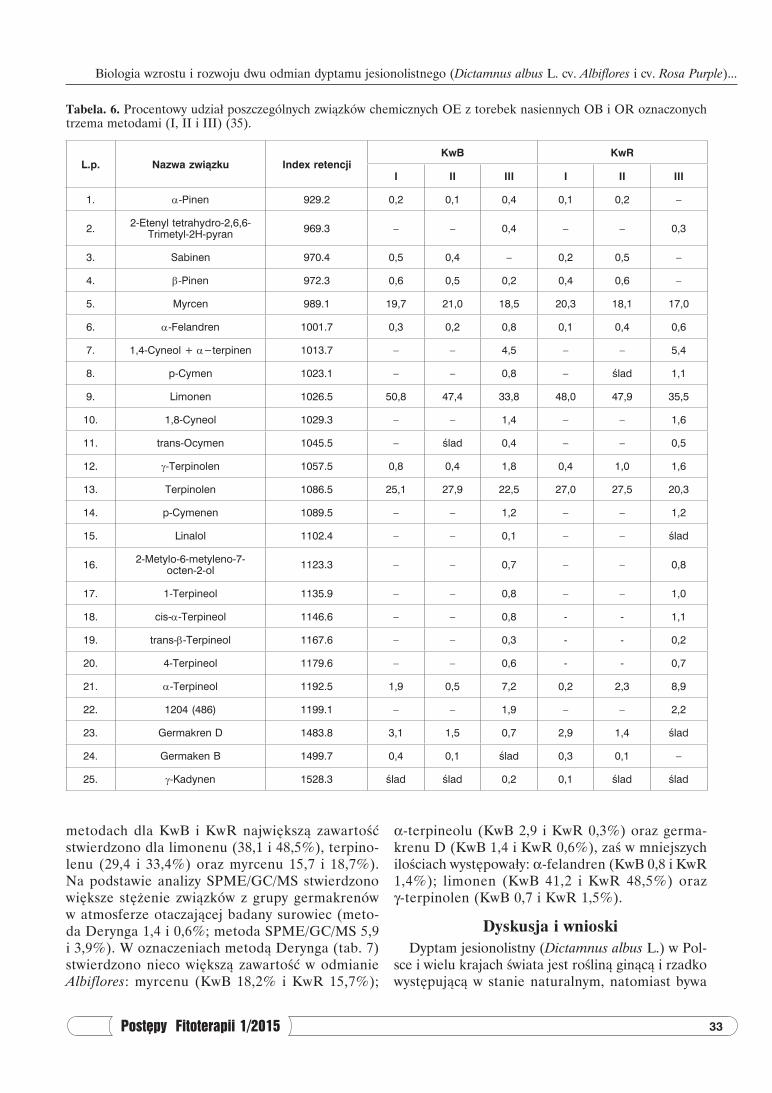
Na podstawie wyników analizy GC/MS olejków eterycznych otrzymanych trzema metodami z owo-costanów obu odmian D. albus, podanych w tabe-li 6, ogółem zidentyfikowano 25 składników. Liczba zidentyfikowanych składników dla OB w metodach I i II wynosiła po 13, zaś w metodzie III – 24; dla OR liczby te przedstawiają się odpowiednio I – 12, II – 13 i III – 21 związków. We wszystkich metodach destylacji z parą wodną (I, II i III) głównymi składni-kami OE w torebkach nasiennych obu odmian były: limonen (dla OB I – 47,7%; II – 46,9%; III – 33,8%, dla OR I – 48,0%; II – 47,9%; III – 35,5%), terpi-nolen (dla OB I – 25,1%; II – 27,9%; III – 22,5%, dla OR I – 27,0%; II – 27,5%; III – 20,3%) i myrcen (dla OB I – 19,7%; II – 21%; III – 18,5%, dla OR I – 20,3%; II – 18,1%; III – 17%).
Jak wynika z przedstawionych danych liczbowych głównymi składnikami OE w torebkach nasiennych obu odmian są monocykliczne terpeny (limonen i ter-pinolen), których procentowy udział mieścił się w gra-nicach 54-76% oraz acykliczny monoterpen (myrcen) stanowiący 17-21% ogólnej sumy olejków.
Kolejnym przedstawicielem zidentyfikowanych monoterpenów był alkohol α-terpineol, którego zawartość, w zależności od metody oznaczania OE, mieściła się w granicach 0,2-8,9%. Oprócz mo-noterpenów, w składzie OE badanych surowców, występowały związki z grupy germakrenów (B i D), których zawartość wynosiła (w zależności od od-miany i stosowanej metody oznaczania) od ilości śladowych do 3,5%.
W metodzie III po hydrolizie kwaśnej zidentyfi-kowano związki, które wcześniej mogły występować w postaci związanej glikozydowo. Wśród związków, których obecności nie stwierdzono (lub ich obecność była śladowa) w poprzednich metodach, w najwięk-szych ilościach oznaczono: 1,4-cyneol+α-terpinen (4,5% – OB, 5,4% – OR), 1,8-cyneol (1,4 i 1,6),
Skład jakościowy i ilościowy olejku eterycznego
Olejek eteryczny (OE) oznaczano w kwiatach oraz torebkach nasiennych. Do oznaczeń OE stosowano trzy metody omówione wcześniej. Do oznaczeń OE w kwiatach stosowano metodę I, zaś w torebkach nasiennych wszystkie trzy metody (I-III). Skład ja-kościowy i ilościowy OE występującego w kwiatach oznaczano metodą GC/MS oraz SPME/GC/MS, zaś w torebkach nasiennych oznaczano metodą GC/MS.
Na rycinie 20 i w tabeli 5 przedstawiono wyniki analizy GC/MS olejków eterycznych występujących w kwiatostanach dwu odmian D. albus i otrzymanych metodą Derynga (metoda I).
W tabeli 6 przedstawiono porównanie składu jako-ściowego i ilościowego olejku eterycznego otrzymane-go trzema metodami (I, II i III) z torebek nasiennych (owocostanów).
Jak wynika z tabeli 5 głównymi składnikami olejku eterycznego w kwiatach dwu odmian D. albus są: limo-
Ryc. 20. Chromatogram analizy GC olejków eterycznych wy-stępujących w kwiatostanach obu odmian D. albus. A – cv. Albiflores, B – cv. Rosa Purple.
A
B
32
Tadeusz Wolski, Tomasz Baj
Postępy Fitoterapii 1/2015
miast wzrost zawartości: α−terpinolenu (7,2% – OB i 8,9% – OR), γ-terpinolenu (odpowiednio 1,8 i 1,6), α-felandrenu (0,8 i 0,6), przy czym nie stwierdzono obecności sabinenu.
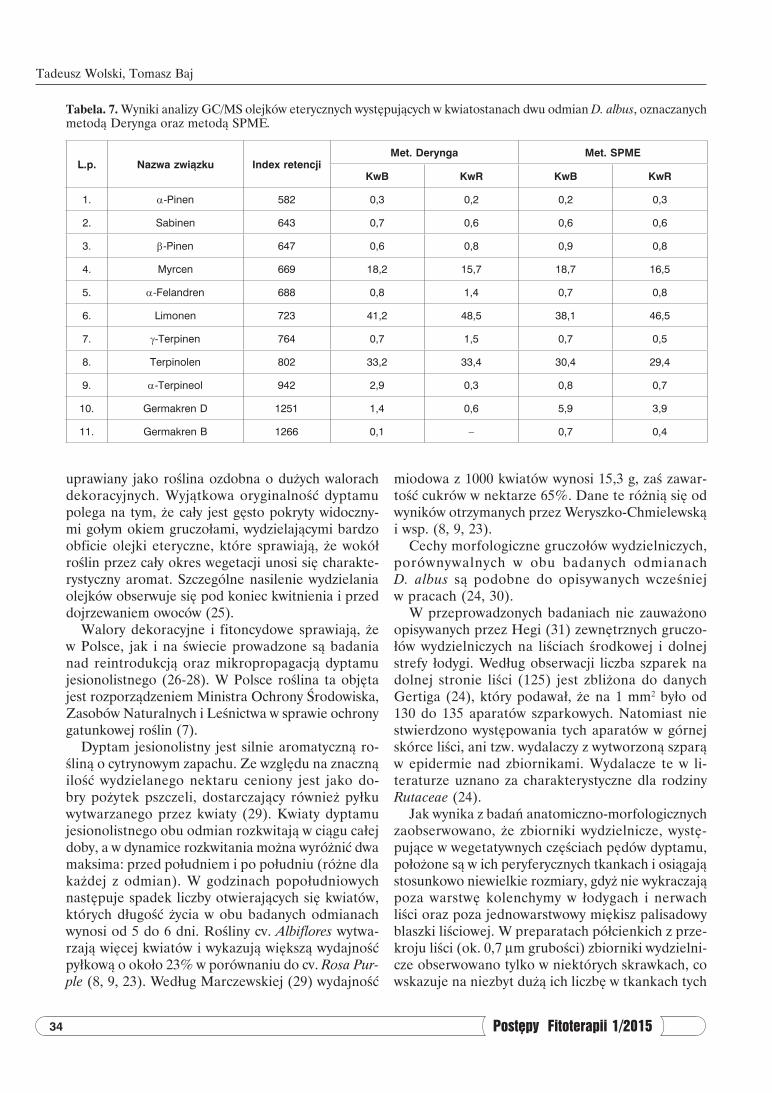
W związku z wprowadzeniem metody sorbcji lotnych składników olejku eterycznego na włóknie adsorpcyjnym w metodzie SPME, postanowiono porównać tę metodę z metodą klasyczną (farma-kopealną). W tabeli 7 przedstawiono wyniki analizy GC/MS olejków eterycznych z kwiatów dwu odmian D. albus oznaczonych metodą destylacji bezpośredniej z parą wodną (met. Derynga) oraz sorbcji na włóknie (metoda SPME). Identyfikację składników olejków eterycznych potwierdzano na podstawie indeksów retencji oraz widm MS.
Jak wynika z tabeli 7 metoda SPME potwierdziła poprzednie badania dotyczące analizy składu jako-ściowego OE dyptamu jesionolistnego. Zawartość głównych składników OE była zbliżona. W obu
p-cymenen (po 1,2%), p-cymen (odpowiednio 0,8 i 1,1) (występował w ilości śladowej w OR, metoda II), cis-β-terpineol (0,8 i 1,1), 1-terpineol (0,8 i 1,0), 2-metylo-6-metyleno-7-octen-2-ol (0,7 i 0,8), 4-terpi-neol (0,6 i 0,7), trans-ocymen (0,4 i 0,5) (występował w ilości śladowej w OB metoda II), 2-etenylo-tetra-hydro-2,6,6-trimetylo-2H-piran (0,4 i 0,3), trans-β-terpineol (0,3 i 0,2), linalol (0,1 – wartość śladowa) (występował w ilości śladowej w OB metoda I); ponadto przy czasie retencji 486 (index 1199.1) wy-stępował związek, którego nie udało się zidentyfiko-wać na podstawie dostępnych wzorców. Z istotnych różnic pomiędzy metodami wymienić należy spadek zawartości limonenu (33,8% – OB. i 35,5% – OR), terpinolenu (odpowiednio 22,5 i 20,3), germakrenu D (0,7 i wartość śladowa), β-pinenu (0,2 i brak) w metodzie III, co prawdopodobnie związane było z ich rozpadem, bądź biotransformacją w środowi-sku kwaśnym. W metodzie III stwierdzono nato-
Tabela. 5. Zawartość procentowa głównych składników OE w KwB i KwR (34).
Lp. Nazwa związku Index retencji KwB Index retencji KwR
1. α-Pinen 596 0,315 599 0,296
2. Kamfen 634 0,081 636 0,072
3. Sabinen 691 0,704 693 0,589
4. β-Pinen 699 0,704 702 0,653
5. Myrcen 736 17,058 737 16,001
6. α-Felandren 771 0,478 773 0,487
7. α-Terpinen 797 0,78 799 0,085
8. p-Cymen 817 0,052 820 0,125
9. Limonen 830 50,127 831 54,313
10. (E)-Ocymen 873 0,110 876 0,087
11. γ-Terpinen 897 0,720 900 0,689
12. Terpinolen 962 23,536 963 22,207
13. Linalool 1010 0,164 1018 0,042
14. (E)-Wodzian pinenu 1059 0,218 1065 0,117
15. (Z)-Wodzian pinenu 1102 0,158 1108 0,042
16. 4-Terpineol 1187 0,173 1194 0,059
17. α-Terpineol 1222 3,147 1229 1,685
18. β-Kariofilen 1689 0,022 1692 0,087
19. α-Humulen 1761 0,009 1763 0,063
20. Germakren D 1811 1,281 1813 2,044
21. Germakren B 1838 0,163 1841 0,257
33
Biologia wzrostu i rozwoju dwu odmian dyptamu jesionolistnego (Dictamnus albus L. cv. Albiflores i cv. Rosa Purple)...
Postępy Fitoterapii 1/2015
metodach dla KwB i KwR największą zawartość stwierdzono dla limonenu (38,1 i 48,5%), terpino-lenu (29,4 i 33,4%) oraz myrcenu 15,7 i 18,7%). Na podstawie analizy SPME/GC/MS stwierdzono większe stężenie związków z grupy germakrenów w atmosferze otaczającej badany surowiec (meto-da Derynga 1,4 i 0,6%; metoda SPME/GC/MS 5,9 i 3,9%). W oznaczeniach metodą Derynga (tab. 7) stwierdzono nieco większą zawartość w odmianie Albiflores: myrcenu (KwB 18,2% i KwR 15,7%);
Tabela. 6. Procentowy udział poszczególnych związków chemicznych OE z torebek nasiennych OB i OR oznaczonych trzema metodami (I, II i III) (35).
L.p. Nazwa związku Index retencjiKwB KwR
I II III I II III
1. α-Pinen 929.2 0,2 0,1 0,4 0,1 0,2 –
2. 2-Etenyl tetrahydro-2,6,6-Trimetyl-2H-pyran 969.3 – – 0,4 – – 0,3
3. Sabinen 970.4 0,5 0,4 – 0,2 0,5 –
4. β-Pinen 972.3 0,6 0,5 0,2 0,4 0,6 –
5. Myrcen 989.1 19,7 21,0 18,5 20,3 18,1 17,0
6. α-Felandren 1001.7 0,3 0,2 0,8 0,1 0,4 0,6
7. 1,4-Cyneol + α−terpinen 1013.7 – – 4,5 – – 5,4
8. p-Cymen 1023.1 – – 0,8 – ślad 1,1
9. Limonen 1026.5 50,8 47,4 33,8 48,0 47,9 35,5
10. 1,8-Cyneol 1029.3 – – 1,4 – – 1,6
11. trans-Ocymen 1045.5 – ślad 0,4 – – 0,5
12. γ-Terpinolen 1057.5 0,8 0,4 1,8 0,4 1,0 1,6
13. Terpinolen 1086.5 25,1 27,9 22,5 27,0 27,5 20,3
14. p-Cymenen 1089.5 – – 1,2 – – 1,2
15. Linalol 1102.4 – – 0,1 – – ślad
16. 2-Metylo-6-metyleno-7-octen-2-ol 1123.3 – – 0,7 – – 0,8
17. 1-Terpineol 1135.9 – – 0,8 – – 1,0
18. cis-α-Terpineol 1146.6 – – 0,8 - - 1,1
19. trans-β-Terpineol 1167.6 – – 0,3 - - 0,2
20. 4-Terpineol 1179.6 – – 0,6 - - 0,7
21. α-Terpineol 1192.5 1,9 0,5 7,2 0,2 2,3 8,9
22. 1204 (486) 1199.1 – – 1,9 – – 2,2
23. Germakren D 1483.8 3,1 1,5 0,7 2,9 1,4 ślad
24. Germaken B 1499.7 0,4 0,1 ślad 0,3 0,1 –
25. γ-Kadynen 1528.3 ślad ślad 0,2 0,1 ślad ślad
α-terpineolu (KwB 2,9 i KwR 0,3%) oraz germa-krenu D (KwB 1,4 i KwR 0,6%), zaś w mniejszych ilościach występowały: α-felandren (KwB 0,8 i KwR 1,4%); limonen (KwB 41,2 i KwR 48,5%) oraz γ-terpinolen (KwB 0,7 i KwR 1,5%).
Dyskusja i wnioskiDyptam jesionolistny (Dictamnus albus L.) w Pol-
sce i wielu krajach świata jest rośliną ginącą i rzadko występującą w stanie naturalnym, natomiast bywa
34
Tadeusz Wolski, Tomasz Baj
Postępy Fitoterapii 1/2015
uprawiany jako roślina ozdobna o dużych walorach dekoracyjnych. Wyjątkowa oryginalność dyptamu polega na tym, że cały jest gęsto pokryty widoczny-mi gołym okiem gruczołami, wydzielającymi bardzo obficie olejki eteryczne, które sprawiają, że wokół roślin przez cały okres wegetacji unosi się charakte-rystyczny aromat. Szczególne nasilenie wydzielania olejków obserwuje się pod koniec kwitnienia i przed dojrzewaniem owoców (25).
Walory dekoracyjne i fitoncydowe sprawiają, że w Polsce, jak i na świecie prowadzone są badania nad reintrodukcją oraz mikropropagacją dyptamu jesionolistnego (26-28). W Polsce roślina ta objęta jest rozporządzeniem Ministra Ochrony Środowiska, Zasobów Naturalnych i Leśnictwa w sprawie ochrony gatunkowej roślin (7).
Dyptam jesionolistny jest silnie aromatyczną ro-śliną o cytrynowym zapachu. Ze względu na znaczną ilość wydzielanego nektaru ceniony jest jako do-bry pożytek pszczeli, dostarczający również pyłku wytwarzanego przez kwiaty (29). Kwiaty dyptamu jesionolistnego obu odmian rozkwitają w ciągu całej doby, a w dynamice rozkwitania można wyróżnić dwa maksima: przed południem i po południu (różne dla każdej z odmian). W godzinach popołudniowych następuje spadek liczby otwierających się kwiatów, których długość życia w obu badanych odmianach wynosi od 5 do 6 dni. Rośliny cv. Albiflores wytwa-rzają więcej kwiatów i wykazują większą wydajność pyłkową o około 23% w porównaniu do cv. Rosa Pur-ple (8, 9, 23). Według Marczewskiej (29) wydajność
Tabela. 7. Wyniki analizy GC/MS olejków eterycznych występujących w kwiatostanach dwu odmian D. albus, oznaczanych metodą Derynga oraz metodą SPME.
L.p. Nazwa związku Index retencjiMet. Derynga Met. SPME
KwB KwR KwB KwR
1. α-Pinen 582 0,3 0,2 0,2 0,3
2. Sabinen 643 0,7 0,6 0,6 0,6
3. β-Pinen 647 0,6 0,8 0,9 0,8
4. Myrcen 669 18,2 15,7 18,7 16,5
5. α-Felandren 688 0,8 1,4 0,7 0,8
6. Limonen 723 41,2 48,5 38,1 46,5
7. γ-Terpinen 764 0,7 1,5 0,7 0,5
8. Terpinolen 802 33,2 33,4 30,4 29,4
9. α-Terpineol 942 2,9 0,3 0,8 0,7
10. Germakren D 1251 1,4 0,6 5,9 3,9
11. Germakren B 1266 0,1 – 0,7 0,4
miodowa z 1000 kwiatów wynosi 15,3 g, zaś zawar-tość cukrów w nektarze 65%. Dane te różnią się od wyników otrzymanych przez Weryszko-Chmielewską i wsp. (8, 9, 23).
Cechy morfologiczne gruczołów wydzielniczych, porównywalnych w obu badanych odmianach D. albus są podobne do opisywanych wcześniej w pracach (24, 30).
W przeprowadzonych badaniach nie zauważono opisywanych przez Hegi (31) zewnętrznych gruczo-łów wydzielniczych na liściach środkowej i dolnej strefy łodygi. Według obserwacji liczba szparek na dolnej stronie liści (125) jest zbliżona do danych Gertiga (24), który podawał, że na 1 mm2 było od 130 do 135 aparatów szparkowych. Natomiast nie stwierdzono występowania tych aparatów w górnej skórce liści, ani tzw. wydalaczy z wytworzoną szparą w epidermie nad zbiornikami. Wydalacze te w li-teraturze uznano za charakterystyczne dla rodziny Rutaceae (24).
Jak wynika z badań anatomiczno-morfologicznych zaobserwowano, że zbiorniki wydzielnicze, wystę-pujące w wegetatywnych częściach pędów dyptamu, położone są w ich peryferycznych tkankach i osiągają stosunkowo niewielkie rozmiary, gdyż nie wykraczają poza warstwę kolenchymy w łodygach i nerwach liści oraz poza jednowarstwowy miękisz palisadowy blaszki liściowej. W preparatach półcienkich z prze-kroju liści (ok. 0,7 µm grubości) zbiorniki wydzielni-cze obserwowano tylko w niektórych skrawkach, co wskazuje na niezbyt dużą ich liczbę w tkankach tych

35
Biologia wzrostu i rozwoju dwu odmian dyptamu jesionolistnego (Dictamnus albus L. cv. Albiflores i cv. Rosa Purple)...
Postępy Fitoterapii 1/2015
Piśmiennictwo1. Baj T. Badania fitochemiczne dwu odmian dyptamu jesiono-
listnego (Disctamnu albus L. cv. Albiflores i cv. Rosa Purple. Roz-prawa doktorska. AM, Lublin 2002; 246. 2. Wolski T, Baj T. Syste-matyka gatunku Dictamnus oraz skład fitochemiczny dwu odmian dyptamu jesionolistnego (Dictamnus albus L. cv. Albiflores i cv. Rosa Purple). Cz. I. Post Fitoter 2014; 3:166-79. 3. Wolski T, Baj T. Systematyka gatunku Dictamnus oraz skład fitochemiczny dwu odmian dyptamu jesionolistnego (Dictamnus albus L. cv. Albiflo-res i cv. Rosa Purple). Cz. II. Post Fitoter 2014; 4:216-26. 4. Ency-klopedia Britanica 2003, www.britanica.com. 5. Wolski T, Werysz-ko-Chmielewska E, Baj T. Zawartość i skład olejków eterycznych dwu odmian dyptamu jesionolistnego (Dictamnus albus L.) cv. Al-biflores i cv. Rosa Purple oraz budowa tkanki wydzielniczej. Anna-les UMCS sec. EEE 1998; 6:151-64. 6. Kostyniuk M, Marczek E. Nasze rośliny chronione. Wrocł Tow Nauk, Wrocław 1961; 82-4:159-60. 7. Rozporządzenie Ministra Środowiska w sprawie ochrony gatunkowej roślin. Dz U 2014; 1409. 8. Weryszko-Chmie-lewska E, Hetman J, Sulborska A i wsp. Ekologia kwiatów i po-żytek pyłkowy dwóch gatunków dyptamu jesionolistnego (Dic-tamnus albus L.), Bibl Fragm Agron 1999; 6:145-51. 9. Weryszko--Chmielewska E., Hetman J., Sulborska A i wsp. Ekologia kwia-tów i pożytek pyłkowy dwóch gatunków dyptamu jesionolistnego (Dictamnus albus L.). Materiały II Ogólnopolsk Konf Nauk nt. Biologia kwitnienia, nektarowania i zapylania roślin. Lublin 1999; 18. 10. Warakomska Z. Badania nad wydajnością pyłkową roślin. Pszczeln Zesz Nauk 1972;16:63:90. 11. Szklanowska K, Pluta S. Wydajność pyłkowa sadu wiśniowego odmian Kerezer, Nefris i Łutówka. Pszczeln Zesz Nauk 1984; 28:163-74. 12. Demiano-wicz Z, Hłyń M, Jabłońska B i wsp. Wydajność miodowa ważniej-szych roślin miododajnych w warunkach Polski (I). Pszczeln Zesz Nauk 1960; 4(2): 87-104. 13. Weryszko-Chmielewska E, Wolski T, Baj T i wsp. Struktura tkanek wydzielniczych w kwiatostanach dyptamu jesionolistnego (Dictamnus albus L. cv. Rosa Purple oraz skład olejku eterycznego. Mat I Ogólnopol Konf Nauk Biologia kwitnienia, nektarowania i zapylania roślin. Lublin 1997; 28-46. 14. Weryszko-Chmielewska E, Baj T, Wolski T. Struktura tkanek wydzielniczych oraz zawartość niektórych substancji biologicznie czynnych w zielu i kwiatach dyptamu jesionolistnego. Herba Pol 1998; 44(4):45-55. 15. Wolski T, Baj T, Weryszko-Chmielewska E. Analiza fitochemiczna ekstraktów z korzeni i ziela oraz budowa tkanki wydzielniczej łodyg i liści dyptamu jesionolistnego (Dic-tamnus albus L. cv. Albiflores), Annales UMCS, sec. EEE 1998; 6:165-84. 16. Wolski T, Baj T, Weryszko-Chmielewska E i wsp. Possibility of photodermatoses arising connected with dittany (Dictamnus albus L.) cultivation. Mat Konf Nauk pt. Ekologickie aspekty v produkcji liecivych rastlin. Nitra (Słowacja) 1998; 56-60. 17. Broda B. Metody histochemii roślinnej. PZWL, Warsza-wa 1971; 245. 18. Farm Polska IV, t II. PZWL, Warszawa 1970. 19. Farm Polska V, t I. PZWL, Warszawa 1990. 20. Farm Polska V, t V. PTFarm, Warszawa 1999. 21. PN-91/R-87019, 1991. Surowce zielarskie. PKNMiJ. 22. Joulian D, Konig WA. The atlas of spec-tral data of sesquiterpene hydrocarbons. E.B. Verlag, Hamburg 1998; 658. 23. Weryszko-Chmielewska E, Masierowska M, Sul-borska A i wsp. Nektarowanie dwóch odmian dyptamu jesiono-listnego (Dictamnus albus L.). Mat 37 Nauk Konf Pszczel, Puławy 2000; 343. 24. Gertig H. Morfologia i anatomia dyptamu jesiono-listnego. Dissert Pharm 1956; 8:203-30. 25. Hetman J, Wolski T. Dyptam roślina nie tylko ozdobna. Kwiaty 1999; 3(103):13-4. 26. Łukasiewicz A. Ekologia, uprawa oraz reintrodukcja Dictam-nus albus L. Prace Ogrodu Bot. PAN 1991; 1:75-85. 27. Jones RO, Geneve RL, Kester ST. Micropropagation of gas plant (Dic-tamnus albus L.). J Environ Hort 1994; 12(4):216-218. 28. Ge-neve RL, Jones RO, Kester ST. Micropropagation of gas plant (Dictamnus albus L.). Springer-Verlag, New York 1997; 360-369.
organów. Potwierdzają to spostrzeżenia dotyczące górnej powierzchni blaszki liściowej, gdzie stwier-dzono również niewielką ich liczbę na powierzchni 1 mm2 (14, 32, 33).
Komórki epidermy występujące nad tkankami sekrecyjnymi łodyg i liści były bardziej spłaszczone i odznaczały się cieńszymi ścianami komórkowymi, niż dalej leżące komórki tej tkanki. Położenie zbior-ników wydzielniczych na brzegach blaszek liściowych i przy ich doosiowej powierzchni wydaje się również sprzyjać efektywniejszemu wydzielaniu olejków ete-rycznych, zwłaszcza w warunkach intensywnej inso-lacji (5, 14, 15).
Obok wykazanych różnic ilościowych, dotyczących wytwarzanego olejku eterycznego przez różne części dyptamu w pracach innych autorów (30), przedstawio-no wyraźne różnice w składzie jakościowym olejków produkowanych przez kwiaty, łodygi i liście wymie-nionego taksonu.
PodsumowanieUprawiane rośliny dyptamu jesionolistnego dobrze
rosły i rozwijały się na poletkach doświadczalnych osiągając wysokość 93 cm dla cv. Albiflores i 110 cm w przypadku cv. Rosa Purple. Kwitnienie obu odmian rozpoczynało się zwykle w trzeciej dekadzie maja i trwało do połowy czerwca. Długość życia kwiatu w zależności od warunków pogodowych trwała ok. 5-7 dni. Kwiaty cv. Albiflores odznaczały się większą obfitością kwitnienia, większą wydajnością pyłkową oraz większą masą wydzielonego nektaru i wydaj-nością miodową w porównaniu z cv. Rosa Purple. Badania morfologiczne i anatomiczne nad strukturą tkanek wydzielniczych obu badanych odmian dyptamu wykazały, że gruczoły i włoski wydzielnicze występują na kwiatach, łodygach i nitkach pręcikowych, i są różnorodne pod względem wielkości, kształtu oraz struktury.
Analiza zawartości i składu olejku eterycznego w badanych surowcach wykazała, że kwiaty: cv. Albiflo-res zawierają 0,76% olejku, zaś cv. Rosa Purple 0,60%; ziele odpowiednio 0,22 i 0,20%; natomiast korzenie 0,10 i 0,17% olejku. Oznaczono również zawartość i skład olejku w torebkach nasiennych trzema meto-dami. Na podstawie tych oznaczeń stwierdzono, że w wyniku hydrolizy kwaśnej uwalniane są glikozydowo związane lotne składniki olejku. Analiza jakościowa i ilościowa metodą GC/MS olejków otrzymanych przez destylację z parą wodną z ziela, kwiatów i korzeni oraz metodą SPME w kwiatach wykazała, że do głównych składników olejków eterycznych obu odmian zaliczyć można: limonen, myrcen, terpinolen, α-terpineol i germakren D.

36
Tadeusz Wolski, Tomasz Baj
Postępy Fitoterapii 1/2015
gruczołów wydzielniczych występujących na powierzchni słupków i owoców dyptamu jesionolistnego (Dictamnus albus L.) Annales UMCS, sec. EEE 2001; 9:339-45. 34. Baj T, Wolski T. Budowa struktur wydzielniczych i skład olejku eterycznego występującego w dyptamie jesionolistnym (Dictamnus albus L.). Aromater 2003; 3-4(9):30-6. 35. Baj T, Wolski T. Ocena zawartości i składu olej-ków eterycznych występujących w owocostanach dwóch odmian dyptamu jesionolistnego (Dictamnus albus L. cv. Albiflores i cv. Rosa Purple). Annales UMCS, sec. EEE 2001; 9:143-50.
29. Marczewska E. Dyptam jesionolistny – chroniona roślina nek-tarodajna. Pszczelnictwo 1976; 4:12-3. 30. Kubeczka KH, Koch V, Ney EM. Das Geheimnis des brennenden Busches. Deuts Apth Ztg 1990; 130(40):2181-5. 31. Hegi G. Illustrierte Flora von Mit-tel Europa. J.I. Lehmanns Verlag, Monachium. 1937; 74-77. 32. Weryszko-Chmielewska E, Baj T, Wolski T. Structure of secre-tory tissues and content of some biologically active compounds in flowers and herb of dittany (Dictamnus albus L.). Mat Konf Nauk Lublin 1998; 52. 33. Weryszko-Chmielewska E. Budowa
otrzymano/received: 10.01.2015zaakceptowano/accepted: 14.02.2015
Adres/address:*prof. dr hab. Tadeusz Wolski
Katedra i Zakład Farmakognozji z Pracownią Roślin Leczniczych
Uniwersytet Medyczny w Lublinie ul. W. Chodźki 1, 20-093 Lublin
tel. +48 (81) 742-38-09e-mail: [email protected]

















![Przywództwo czynnikiem rozwoju i wzrostu innowacyjnej ...€¦ · B.M. Bassa [1985]. Na podstawie analizy literaturowej scharakteryzowano mechanizmy, dzięki którym przywódcy mogą](https://static.fdocument.pub/doc/165x107/5f9aa77b7fb5340af55041d8/przywdztwo-czynnikiem-rozwoju-i-wzrostu-innowacyjnej-bm-bassa-1985-na.jpg)
