
Aus dem Institut für Zelluläre und Molekulare Physiologie ... · Ami) mit der...
77
Aus dem Institut für Zelluläre und Molekulare Physiologie der Friedrich-Alexander-Universität Erlangen-Nürnberg Direktor: Prof. Dr. med. Christoph Korbmacher Funktionelle Charakterisierung der βV348M-Mutation des epithelialen Natriumkanals (ENaC) Inaugural-Dissertation zur Erlangung der Doktorwürde der Medizinischen Fakultät der Friedrich-Alexander-Universität Erlangen-Nürnberg vorgelegt von Daniel Söll aus Bayreuth
Transcript of Aus dem Institut für Zelluläre und Molekulare Physiologie ... · Ami) mit der...

Aus dem Institut für Zelluläre und Molekulare Physiologie
der
Friedrich-Alexander-Universität Erlangen-Nürnberg
Direktor: Prof. Dr. med. Christoph Korbmacher
Funktionelle Charakterisierung der βV348M-Mutation des
epithelialen Natriumkanals (ENaC)
Inaugural-Dissertation zur
Erlangung der Doktorwürde
der Medizinischen Fakultät
der
Friedrich-Alexander-Universität
Erlangen-Nürnberg
vorgelegt von
Daniel Söll
aus
Bayreuth

Gedruckt mit Erlaubnis der Medizinischen Fakultät der
Friedrich-Alexander-Universität Erlangen-Nürnberg
Dekan: Prof. Dr. med. Dr. h. c. Jürgen Schüttler
Referent: Priv.-Doz. Dr. med. Robert Rauh
Korreferent: Prof. Dr. med. Christoph Korbmacher
Tag der mündlichen Prüfung: 05. Juni 2013

Meiner Familie

Inhalt
1. Zusammenfassung 1
1.1 Hintergrund und Ziele 1
1.2 Methoden 1
1.3 Ergebnisse 1
1.4 Praktische Schlussfolgerungen 2
2. Summary 3
2.1 Background and Aim of the Study 3
2.2 Methods 3
2.3 Results 3
2.4 Conclusion 3
3. Einleitung 4
3.1 Zystische Fibrose (CF) 4
3.2 Struktur von ENaC 5
3.3 Lokalisation und Funktion von ENaC 7
3.4 Pathophysiologie des Lungenphänotyps bei CF 10
3.5 Zielsetzung der Arbeit: funktionelle Charakterisierung der βV348M-
Mutation 12
4. Material und Methoden 13
4.1 Xenopus laevis Oozyten als heterologes Expressionssystem 13
4.2 Lösungen und Chemikalien 15
4.3 cRNA-Herstellung 16
4.4 Isolation und Injektion der Oozyten 16
4.5 Elektrophysiologische Methoden 18
4.5.1 Zwei-Elektroden-Spannungsklemme 18
4.5.2 Messablauf 19
4.6 Proteinnachweis 20
4.6.1 Nachweis membranständiger Proteine durch Biotinylierung 20
4.6.2 Proteinnachweis durch Western Blot 22
4.7 Statistik 23

5. Ergebnisse 24
5.1 Die βV348M-Mutation stimuliert den ENaC-vermittelten Na+-Strom 24
5.2 Der stimulatorische Effekt der βV348M-Mutation ist vom Na+-Gehalt
der Oozyten weitgehend unabhängig 26
5.3 Die βV348M-Mutation erhöht die mittlere Po von ENaC 27
5.4 Chymotrypsin stimuliert αβγENaC mehr als αβV348MγENaC 30
5.5 Die βV348M-Mutante wird durch S3969 weniger aktiviert als der
Wildtyp 32
5.7 Verschiedene Mutationen an βV348 36
5.7.1 Aminosäure-Mutationen zeigen unterschiedliche Effekte auf die ENaC-Funktion 36
5.7.2 Effekte der Mutationen auf die Proteinexpression 38
5.7.3 Proteolytische Aktivierung der mutierten Kanäle mit Chymotrypsin 41
6. Diskussion 43
6.1 Xenopus laevis Oozyten als heterologes Expressionssystem 43
6.2 Einflüsse der cRNA-Qualität 44
6.3 Einfluss der intrazellulären Na+-Konzentration 45
6.4 Mittlere Po 45
6.5 Proteolytische Kanalaktivierung mit Chymotrypsin 47
6.6 ENaC-Aktivator S3969 48
6.7 Weitere βV348-Mutanten und ENaC-Modell 49
6.8 Klinische Relevanz der βV348M-Mutation 52
7. Schlussfolgerung 54
8. Literaturverzeichnis 55
9. Abbildungsverzeichnis 62
10. Abkürzungsverzeichnis 63
10.1 Abkürzungen 63
10.2 Nomenklatur der Aminosäuren 65
10.3 Einheiten 65
11. Vorveröffentlichungen 66
12. Anhang 67
13. Danksagung 68
14. Lebenslauf 70

1
1. Zusammenfassung
1.1 Hintergrund und Ziele
Bei einigen Patienten mit atypischer zystischer Fibrose (CF) ist nur ein oder
kein Allel des „cystic fibrosis transmembrane conductance regulator“ (CFTR)-
Gens mutiert, das einen Chloridkanal kodiert. Mutationen des epithelialen Na+-
Kanals (ENaC) könnten zur Pathophysiologie der CF in diesen Patienten
beitragen. Das Ziel dieser Arbeit war die funktionelle Charakterisierung einer
Mutation in der β-Untereinheit von ENaC (βV348M), die kürzlich in einem
Patienten mit CF-ähnlichen Symptomen identifiziert wurde.
1.2 Methoden
αβγENaC (Wildtyp) und αβV348MγENaC (βV348M-Mutante) wurden in Xenopus
laevis Oozyten als heterologem Expressionssystem exprimiert. Die ENaC-
Aktivität wurde durch die Messung des Amilorid-sensitiven Ganzzellstroms
(ΔIAmi) mit der Zwei-Elektroden-Spannungsklemme bestimmt. Die Expression
von ENaC in der Zellmembran wurde durch Biotinylierung, Western Blot und
densitometrische Auswertung der Signale untersucht.
1.3 Ergebnisse
ΔIAmi erwies sich in αβV348MγENaC exprimierenden Oozyten um durchschnittlich
~40% höher als in αβγENaC exprimierenden Oozyten. Während die Expression
in der Zellmembran von Wildtyp und Mutante ähnlich war, zeigte sich eine
erhöhte mittlere Offenwahrscheinlichkeit (Po) des Wildtyps gegenüber der
Mutante (~0,33 vs. ~0,24). Diesem Ergebnis entsprechend war die
proteolytische Aktivierung durch Chymotrypsin im mutierten Kanal gegenüber
dem Wildtyp reduziert (~3-fach vs. ~4,2-fach). Der stimulatorische Effekt des
ENaC-Aktivators S3969 auf den mutierten Kanal war im Vergleich zum Wildtyp
in ähnlicher Weise reduziert (~2,6-fach vs. ~3,5-fach). Die funktionelle
Expression verschiedener βV348-Mutanten in Oozyten in Verbindung mit der
Computer-gestützten Analyse eines virtuellen ENaC-Modells lässt vermuten,
dass die βV348M-Mutation die Po des Kanals durch die Destabilisierung des
geschlossenen Kanal-Zustands erhöht.

2
1.4 Praktische Schlussfolgerungen
Die Ergebnisse weisen darauf hin, dass der funktionssteigernde Effekt der
βV348M-Mutation zu einer unangemessen hohen Resorption von Natrium und
Flüssigkeit im Respirationstrakt führt und dadurch zur Pathophysiologie der CF-
Symptomatik bei betroffenen Patienten beiträgt.

3
2. Summary
2.1 Background and Aim of the Study
In some patients with atypical cystic fibrosis (CF) only one or no allele of the
cystic fibrosis transmembrane conductance regulator (CFTR) gene is affected
which codes for a chloride channel. Mutations of the epithelial sodium channel
(ENaC) may contribute to the pathophysiology in these patients. The aim of the
present study was to functionally characterize a mutation in the β-subunit of
ENaC (βV348M) recently identified in a patient with severe CF-like symptoms.
2.2 Methods
Wild-type (wt) αβγENaC or mutant αβV348MγENaC was expressed in Xenopus
laevis oocytes and ENaC function was assessed by measuring the amiloride
sensitive whole-cell current (ΔIami) with two-electrode voltage-clamp. ENaC cell
surface expression was evaluated by biotinylation, Western Blot and
subsequent densitometric analysis of the signals.
2.3 Results
ΔIami was on average ~40% higher in mutant ENaC than in wt ENaC expressing
oocytes. Whereas surface expression of wt and mutant ENaC were similar,
average channel open probability (Po) of wt ENaC was higher than that of
mutant ENaC (~0.33 vs. ~0.24). In agreement with that result, proteolytic
activation by chymotrypsin was reduced in mutant ENaC compared to that of wt
ENaC (~3fold vs. ~4.2fold). Similarly, the ENaC activator S3969 stimulated
mutant ENaC currents to a lesser degree than wt ENaC currents (~2.6fold vs.
~3.5fold). Functional expression of different βV348 mutants in oocytes in
combination with computational channel modeling suggests that the βV348M
mutation increases channel Po by destabilizing the closed channel state.
2.4 Conclusion
The findings indicate that the gain-of-function effect of the βV348M mutation
may contribute to CF pathophysiology by inappropriately increasing sodium and
fluid absorption in the respiratory tract.

4
3. Einleitung
3.1 Zystische Fibrose (CF)
Die Zystische Fibrose (Mukoviszidose) stellt die häufigste letale genetische
Erkrankung der hellhäutigen Bevölkerung dar. Sie wird autosomal rezessiv
vererbt und beruht im Regelfall auf zwei mutierten Allelen des CFTR-Gens [39,
57]. Das CFTR-Gen kodiert für einen Cl--Kanal in diversen Epithelien [63].
Mutierte CFTR-Kanäle verursachen letzten Endes eine geringere Leitfähigkeit
für Cl-, was den transepithelialen Chloridtransport beeinträchtigt. Dies hat in
sekretorischen Epithelien zur Folge, dass die epithelialen Sekrete eine erhöhte
Viskosität aufweisen, was zu den Symptomen der CF beiträgt, die auch als
Mukoviszidose bezeichnet wird.
Zu den pulmonalen Symptomen der CF zählen chronische Bronchitiden und
Pneumonien mit irreversiblem Umbau des Lungengerüsts. Dieser Umbau wird
durch die chronische Besiedlung des zähen Schleims in der Lunge mit
Pseudomonas aeruginosa und/oder Staphylococcus aureus verursacht. Zum
CF-Symptomkomplex gehören auch gastrointestinale Symptome wie ein
Mekoniumileus, Darmobstruktionen, Diarrhö, Obstipation und Leber- und
Gallenwegsdysfunktionen (Leberzirrhose, Gallensteine). Außerdem kommt es
häufig zur exokrinen aber auch endokrinen Pankreasinsuffizienz, was zur
Proteinmangelernährung, Gedeihstörung (auch durch die chronische
Lungenaffektion verstärkt) und Diabetes mellitus führen kann. Auch eine durch
abnorme Sekretbildung in den Samenwegen verursachte Infertilität bei
männlichen Patienten zeigt eine hohe Prävalenz [52]. Hierbei handelt es sich
um ein breites Symptomspektrum, dass von Patient zu Patient sehr heterogen
ausgeprägt ist. Es gibt Patienten, die fast alle klassischen Symptome
aufweisen, während andere nur eine sehr milde Verlaufsform zeigen [18].
In den Ausführungsgängen der Schweißdrüsen führt eine gestörte CFTR-
Funktion zu einer verminderten Chloridresorption und dadurch zu einer
erhöhten Chloridkonzentration im Schweiß. Die Bestimmung der
Chloridkonzentration im Schweiß gehört daher zur Diagnostik bei Verdacht auf
CF. Man unterscheidet zwei unterschiedliche Entitäten der CF. Bei einer
Chlorid-Konzentration über 60 mmol/l spricht man von klassischer bzw.
typischer CF, bei einer Konzentration von 30-60 mmol/l (Borderline) oder sogar

5
unter 30 mmol/l (Normalbereich) von nicht-klassischer oder atypischer CF,
wenn gleichzeitig in mindestens einem Organsystem für die CF typische
Symptome auftreten. Die Einteilung in diese Kategorien gestaltet sich aber
aufgrund des Kontinuums an Symptomen schwierig [18]. Die Diagnose CF wird
mit Tests auf Mutationen im CFTR-Gen gesichert [18].
Die große Heterogenität an Symptomen und eine schwache Genotyp-Phänotyp
Beziehung ließen die Existenz von modifizierenden Genen („Modifier genes“) in
der Pathogenese der CF vermuten [17]. So konnten die für die β- und γ-
Untereinheit des epithelialen Natrium Kanals (ENaC) kodierenden Gene als
potentielle „Modifier“-Gene identifiziert werden [67].
Typische CF-Symptome können auch dann auftreten, wenn im CFTR-Gen kein
oder nur ein Allel eine Mutation aufweist [18, 51]. In manchen Patienten mit
einer milden Ausprägung der Krankheit wurde eine solche Konstellation der
Gene gefunden [3, 64].
Dies führte zu der Fragestellung, ob diese Patienten Mutationen in den für
ENaC kodierenden Genen aufwiesen. Tatsächlich konnten bei Patienten mit
atypischer CF und keinem [3, 64] oder nur einem [3] betroffenen CFTR-Allel
Mutationen in den für ENaC kodierenden Genen identifiziert werden.
3.2 Struktur von ENaC
Der epitheliale Natriumkanal (ENaC) gehört zur Familie der nicht
spannungsabhängigen ENaC/Degenerin-Ionenkanäle [38]. Er besteht aus den
drei Untereinheiten α, β und γ [38, 56]. Die Homologien der Aminosäurenketten
der Untereinheiten belaufen sich auf ca. 30-40% [11, 38, 56]. Strukturell bilden
die drei Untereinheiten vermutlich ein Heterotrimer in der Zellmembran aus
(Abb. 1). Das lässt sich von der erst veröffentlichten Kristallstruktur des
verwandten „acid sensing Ion channel 1“ (ASIC 1) ableiten [10, 35, 68]. Die
Untereinheiten besitzen eine Länge von 669 (α), 640 (β) bzw. 649 (γ)
Aminosäuren und gliedern sich in jeweils zwei Transmembrandomänen, eine
ausgedehnte extrazelluläre Schleife und jeweils einen intrazellulär gelegenen
Amino (N)- und Carboxy (C)-Terminus [38]. Interessanterweise existiert beim
Menschen im Gegensatz zu Maus und Ratte eine zusätzliche δ-Untereinheit,
deren Funktion jedoch noch weitestgehend ungeklärt ist [30, 73].

6
Abb. 1: Struktur von ENaC in der Zellmembran. ENaC ist als Heterotrimer, bestehend aus α-
(rot), β- (gelb), und γ-Untereinheit (blau), dargestellt. M1 bzw. M2 kennzeichnen die
Transmembrandomänen, N und C entsprechend Amino(N)- bzw.- Carboxy(C)-Terminus der
Untereinheiten.
Der Versuch, ENaC zu kristallisieren, gelang bis heute nicht, so dass dessen
Struktur noch nicht endgültig geklärt werden konnte.
Auf Homologie basierende Modellberechnungen, die sich an der bekannten
Kristallstruktur des aus der gleichen Ionenkanal-Familie wie ENaC stammenden
ASIC 1 Ionenkanals orientieren [35, 68], liefern die bisher beste molekulare
Darstellung der Kanalstruktur von ENaC.
Für die vorliegende Arbeit wurde ein entsprechendes Homologie-Modell von
Prof. Heinrich Sticht (Abteilung für Bioinformatik, Institut für Biochemie,
Universität Erlangen-Nürnberg) entworfen, das den Kanal vermutlich im
geschlossenen Zustand darstellt. In diesen Berechnungen wurde insbesondere
die molekulare Mikrodomäne in der Umgebung der Aminosäure V348 der β-
Untereinheit analysiert, die im Fokus dieser Arbeit steht. Die strukturelle
Analyse des aus einem Trimer der Untereinheiten α, β und γ bestehenden
ENaC-Modell lässt vermuten, dass V348 der β-Untereinheit in einer
kugelförmigen Domäne, der sogenannten „palm“-Domäne liegt. Die „Palm“-
Region befindet sich in der Nähe der Zellmembran (Abb. 2).

7
Abb. 2: ENaC-Modell (mod. nach [55]). A, Trimer aus den drei Untereinheiten α, β und γ in
der geschlossenen Konformation. Die β-Untereinheit ist blau gefärbt, die Aminosäuren V348
und M90 sind im Kalottenmodell dargestellt. B, Struktur der β-Untereinheit. Der Aufbau in
Domänen ist farblich hervorgehoben. Die verschiedenen ENaC-Domänen sind entsprechend
der von Stockand et al. [68] verwendeten Terminologie benannt und folgendermaßen gefärbt:
Transmembran-Domäne (rot), „Palm“-Domäne (gelb), „β-Ball“-Domäne (braun), „Finger“-
Domäne (Magenta), „Knuckle“-Domäne (cyan), und „Thumb“-Domäne (grün).
3.3 Lokalisation und Funktion von ENaC
ENaC wird in vielen unterschiedlichen Geweben des menschlichen Körpers
exprimiert. So lässt sich zum Beispiel in Epithelzellen Na+-resorbierender
Gewebe eine starke Expression von ENaC nachweisen, wie im distalen Kolon
[20, 25] oder in Schweiß- [61] und Speicheldrüsen [20].
ENaC wird außerdem in der apikalen Membran der Tubuluszellen des
Aldosteron-sensitiven distalen Nephron (ASDN) der Niere exprimiert. Das
ASDN umfasst den Endabschnitt der Pars convoluta des distalen Tubulus, den
Verbindungstubulus und das Sammelrohr. Dort ist ENaC entscheidend am
transepithelialen Na+-Transport beteiligt (Abb. 3). Voraussetzung für die Na+-
Resorption ist der primär aktive Transport von Na+-Ionen über die basolaterale
Membran ins renale Interstitium durch die Na+/K+-ATPase. Im Gegenzug
transportiert die Na+/K+-ATPase K+-Ionen aus dem Interstitium in die Zellen.
A B

8
ROMK
Abb. 3: Von ENaC vermittelter Na+-Transport durch eine Hauptzelle im ASDN.
Die K+-Ionen rezirkulieren aus der Zelle ins Interstitium über basolaterale
Kaliumkanäle, wodurch ein negatives Membranpotential aufrecht erhalten wird.
Die niedrige intrazelluläre Na+-Konzentration und das negative
Membranpotential generieren den elektrochemischen Gradienten für den
ENaC-vermittelten Einstrom der Na+-Ionen aus dem Tubuluslumen in die
Tubuluszelle. Dabei ist die Aktivität von ENaC ratenlimitierend für die Na+-
Resorption und ist daher von entscheidender Bedeutung für die Feinregulation
des Na+-Haushaltes und des extrazellulären Flüssigkeitsvolumens und somit für
die langfristige Einstellung des arteriellen Blutdrucks [1, 28, 38, 59]. Aufgrund
des Na+-Einstroms depolarisiert die apikale Membran, was eine „Lumen-
negative“ transepitheliale Potentialdifferenz zur Folge hat, die umso negativer
ist, je aktiver ENaC ist. Diese transepitheliale Potentialdifferenz fördert die
parazelluläre Resorption von Cl- Ionen und die Sekretion von K+-Ionen über
apikale Kaliumkanäle vom Typ des „renal outer medullary“-Kaliumkanals
(ROMK) [50, 75]. Im Endeffekt kommt es bei einer entsprechenden Aktivität von
ENaC zu einer Resorption von Na+- und Cl--Ionen und bei entsprechender
Wasserpermeabilität aufgrund der osmotischen Triebkraft auch zu einer
Wasserresorption [38].

9
Die physiologische Bedeutung von ENaC in der Niere wird durch zwei
genetische Erkrankungen unterstrichen, das Liddle-Syndrom und den
Pseudohypoaldosteronismus Typ I (PHA 1). Die Ursachen dieser
Fehlfunktionen liegen jeweils in Mutationen in einem der für die ENaC-
Untereinheiten kodierenden Gene. Die Erkrankungen sind zwar selten, zeigen
aber direkt, dass molekulare Dysfunktionen in ENaC gravierende
Krankheitssymptome nach sich ziehen können.
Das Liddle-Syndrom stellt eine Form der sekundären arteriellen Hypertonie dar,
welche ursächlich auf einer funktionssteigernden Mutation, einer sog. “gain-of-
function“-Mutation, in ENaC beruht und dadurch zu gesteigerter Na+- und
Wasser-Resorption führt. Auch eine Hypokaliämie und metabolische Alkalose
ist bei den betroffenen Patienten nachzuweisen [65].
Der PHA I wird dagegen durch funktionseinschränkende Mutationen, sog. “loss-
of-function“-Mutationen, in ENaC ausgelöst und führt dadurch zu beträchtlichem
renalen Salzverlust. Dieser geht mit einer Hyperkaliämie, metabolischer
Azidose und arterieller Hypotension einher [13, 38].
Im respiratorischen Epithel ist ENaC ebenfalls von entscheidender Bedeutung
für die Salz- und Flüssigkeitsresorption [7, 26, 34, 70]. ENaC reguliert dadurch
die Höhe und Viskosität des das Lungenepithel bedeckenden Flüssigkeitsfilms,
der sogenannten „Airway surface liquid“ (ASL). Die ASL besteht aus einer die
Zilien umgebenden flüssigen bzw. gelartigen Schicht, dem sogenannten
„periciliary layer“ (PCL), und dem darauf schwimmenden Mukus. Diese
Schleimschicht schützt vor Austrocknung. Außerdem werden in ihr inhalierte
Partikel und Bakterien „gefangen“. Diese werden dann zusammen mit dem
Schleim von den Zilien des Respirationsepithels zum Pharynx transportiert, wo
der Schleim durch Husten oder Schlucken aus dem Respirationstrakt entfernt
wird. Diesen Prozess nennt man mukoziliäre Clearance und er sorgt für die
Reinigung von inhalierten Partikeln [8].
Das Volumen der ASL ist für die mukoziliäre Clearance entscheidend. Es wird
von CFTR und ENaC in entgegengesetzter Weise reguliert (Abb. 4). Die Cl--
Sekretion mittels CFTR und in geringerem Ausmaß mittels auswärts
gleichrichtenden Ca2+-aktvierten Cl--Kanälen erhöht die Osmolarität der ASL
und fördert dadurch die Flüssigkeitssekretion. ENaC wirkt diesem Effekt von
CFTR entgegen, indem mittels Na+-Resorption und osmotisch folgender
Wasser-Resorption die Höhe der ASL gesenkt wird [6, 12].

10
Abb. 4: Regulation und Fehlregulation der ASL. Links ist die physiologische Situation mit
normaler CFTR- und ENaC-Funktion dargestellt. Die für die Ausprägung des CF-
Lungenphänotyps typische Verringerung der Höhe der ASL kann auf einer CFTR-Unterfunktion
(Mitte) oder auf einer ENaC-Überfunktion (rechts) beruhen.
3.4 Pathophysiologie des Lungenphänotyps bei CF
Die exakte Regulation der Höhe der ASL ist eine wichtige Voraussetzung für
eine angemessene Abwehrfunktion der Lunge. Die Abnahme des
Wassergehalts der ASL führt zur Reduktion der Höhe der ASL und zur Bildung
eines visköseren Mukus. Zusätzlich wird die Zilienbewegung behindert. Die
Folge ist eine Verschlechterung der mukoziliären Clearance [42], wodurch die
chronische Besiedlung mit pathogenen Keimen begünstigt wird, was zur
Pathophysiologie der pulmonalen Symptome bei Patienten mit zystischer
Fibrose (CF) maßgeblich beiträgt. So wurde in Zellkulturen von menschlichen
Epithelzellen des Respirationstraktes von gesunden Spendern eine Höhe des
PCL von etwa 7 µm nachgewiesen, was interessanterweise der Höhe von
„gestreckten“ Zilien entspricht. Der PCL von Epithelzellen von CF-Patienten war
dagegen stark reduziert (~3 µm) [49].
Am häufigsten liegt der Ausbildung von pulmonalen Symptomen bei CF eine
Verringerung der CFTR-Funktion zugrunde (Abb. 4). Defekte CFTR-Kanäle
reduzieren die Cl--Sekretion und dadurch die Höhe der ASL. Seit der
erfolgreichen Klonierung von CFTR und dem Nachweis, dass CFTR die
Funktion eines Chloridkanals hat, wird CF als Krankheit angesehen, die
hinsichtlich der pulmonalen Symptomatik vor allem auf einer gestörten Cl--
Sekretion beruht.

11
Nach neueren Erkenntnissen kann aber auch eine erhöhte ENaC-Funktion zur
pulmonalen Pathophysiologie von CF beitragen [5, 6]. Eine vermehrte ENaC-
Aktivität im respiratorischen Epithel führt durch gesteigerte Na+- und
Flüssigkeitsresorption zur Verringerung der ASL (Abb. 4). So zeigten Mall et al.
[47] in transgenen Mäusen, in denen eine Überexpression der β-Untereinheit
von ENaC induziert wurde, dass eine Überfunktion von ENaC zu einer CF-
ähnlichen Lungenaffektion mit verminderter ASL und fehlerhaftem Mukus-
Transport führt. Interessanterweise konnte in diesen Mäusen durch präventive
Behandlung mit Amilorid die Morbidität und Mortalität gesenkt werden [76].
Diesem Effekt liegt vermutlich die Hemmung von ENaC durch Amilorid, einem
spezifischem ENaC-Inhibitor (Abb. 3), zugrunde. Ganz ähnliche Effekte wurden
auch in Knock-out Mäusen für die Ubiquitin-Ligase Nedd4-2 entdeckt, die
physiologischerweise ENaC degradiert. Eine gesteigerte ENaC-Expression war
auch hier die Ursache für die Ausbildung einer CF-ähnlichen Lungenaffektion,
die ebenfalls durch präventive Amilorid-Applikation gemildert werden konnte
[41]. Diese Befunde unterstützen die Hypothese, dass auch „gain-of-function“-
Mutationen von ENaC zur Pathophysiologie der CF beitragen können.
Von Mutesa et al. wurde in Ruanda, wo die CF aufgrund der Ähnlichkeit der
Symptome zu anderen häufigen Krankheitsbildern wie der Malnutrition eher
unterdiagnostiziert wird, eine Genanalyse der für die verschiedenen ENaC-
Untereinheiten kodierenden Sequenzen durchgeführt [51]. Die Genanalyse von
Mutesa et al. umfasste 60 Kinder, die alle an CF-ähnliche Symptomen litten,
aber nur ein oder gar kein mutiertes CFTR-Allel aufwiesen. Bei den Patienten,
die nur ein mutiertes CFTR-Allel besaßen, wurden mehrere
Sequenzänderungen in den Genen der ENaC-Untereinheiten detektiert. Dabei
fiel vor allem eine Missense-Mutation bei einem 13-jährigen Patienten auf. Sein
CF-Phänotyp zeigte sich vor allem in einer ausgeprägten Lungenbeteiligung im
Sinne der Besiedelung mit Pseudomonas aeruginosa und rezidivierenden
respiratorischen Infektionen. Die Krankheit manifestierte sich bei ihm auch in
Form von gastrointestinalen Symptomen, Gedeihstörung, Diabetes mellitus und
Proteinmangelernährung. Der Schweißtest war mit einer Chloridkonzentration
von 94 mmol/l deutlich im pathologischen Bereich, was für eine typische CF
spricht. Die genetische Konstellation mit nur einem betroffenen CFTR-Allel stellt
sich jedoch als atypisch dar.

12
3.5 Zielsetzung der Arbeit: funktionelle Charakterisierung der
βV348M-Mutation
Die Missense-Mutation des Patienten wurde in einer hoch konservierten Region
der extrazellulären Schleife der β-Untereinheit von ENaC an Position 348
gefunden (Abb. 2 und 5), an der Valin zu Methionin mutiert war. Die βV348M-
Mutation wurde in keinem anderen Patienten und auch keiner Person des
Kontrollkollektivs gefunden, so dass eine funktionelle Relevanz wahrscheinlich
ist.
Da bei dem Patienten nur eine Mutation in einem Allel des CFTR-Gens
nachgewiesen wurde, aber schwere Symptome der CF auftraten, war es Ziel
dieser Arbeit, die bei ihm nachgewiesene βV348M-Mutation von ENaC im
Oozytenexpressionsmodell funktionell zu charakterisieren.
Abb. 5: Aminosäuresequenzen verschiedener Spezies der Region um βV348. Die Zahlen
geben Beginn und Ende des dargestellten Ausschnittes der Aminosäurekette der β-Untereinheit
beginnend vom N-Terminus an.
Mensch 341 ETSIGVLVDKLQRMG 355
Maus 339 ETSIGVLVDKLQRKG 353
Ratte 339 ETSIGVLVDKLQRKG 353
Meerschweinchen 342 ETSIGVLVDKLQRMG 356
Kaninchen 342 ETSIGVLVDKLQRKG 356
Riesenpanda 347 ETSIGVLVDKLERKG 361
Rind 342 ETSIGVLVDKLQRKG 356
Xenopus laevis 361 ETSISVLVDQLEHME 375
Xenopus tropicalis 360 ETSIAVLVDQLEHME 374

13
4. Material und Methoden
4.1 Xenopus laevis Oozyten als heterologes Expressions-
system
In früheren Jahren wurde der afrikanische Krallenfrosch Xenopus laevis (Abb.
6) als Versuchstier zum Schwangerschaftsnachweis verwendet. Dazu wurde
den Weibchen der Urin von fraglich schwangeren Frauen injiziert. Eine Eiablage
der Weibchen durch im Urin enthaltenes humanes Choriongonadotropin
kennzeichnete den positiven Test [24]. Da dieser Test in den 40er Jahren in
Apotheken zum Schwangerschaftsnachweis durchgeführt wurde, wurde der
Frosch als „Apothekerfrosch“ bekannt. Heute werden vor allem die Oozyten von
Xenopus laevis in der Forschung verwendet.
Gurdon et al. [29] konnten 1971 zum ersten Mal zeigen, dass injizierte exogene
mRNA in den Oozyten die Bildung der dafür kodierenden Proteine im
Zytoplasma der Oozyte zur Folge hat. Hierauf konnten etliche Signalkaskaden
mit Hilfe dieses Systems analysiert werden. Mit elektrophysiologischen
Methoden wurden schließlich ligandengesteuerte und spannungsabhängige
Ionenkanäle untersucht [4, 69].
Dabei eignet sich das Expressionssystem zur systematischen Untersuchung
der Beziehung von Funktion und Struktur der Ionenkanäle durch den Vergleich
von Wildtyp-Proteinen mit verschiedenen durch positionsgerichtete Mutagenese
hergestellten Mutanten [16].
Abb. 6: Xenopus laevis, der afrikanische Krallenfrosch.

14
Die Entwicklungsstufen der Oozyten werden nach Dumont in sechs Stadien
eingeteilt [21]. Diese unterscheiden sich anhand der Größe und Pigmentierung
der Oozyten, wobei in der vorliegenden Arbeit nur die Stadien V und VI für die
Messungen herangezogen wurden. In Stadium V und VI ist die morphologische
Zweiteilung in einen animalen und einen vegetativen Pol gut erkennbar (Abb.
7). Die vegetative Hemisphäre ist dabei hell gefärbt, während die animale
Hemisphäre durch eingelagertes Melanin dunkel gefärbt ist [16]. Bei Oozyten
des Stadiums VI fällt ein nicht pigmentierter Äquatorialring auf, der die beiden
Pole voneinander trennt.
Abb. 7: Xenopus laevis Oozyten im Reifestadium V und VI nach Dumont.

15
4.2 Lösungen und Chemikalien
Die in der vorliegenden Arbeit verwendeten Lösungen sind in Tabelle 1 und 2
aufgeführt. Die Lösungen wurden vor Verwendung frisch hergestellt. ND96 und
OR2 wurden aus einer autoklavierten 10-fach Stammlösung zubereitet.
Tabelle 1: Lösungen für Präparation und Inkubation der Oozyten
OR2 ND9 ND96
NaCl (mM) 82,5 9 96
KCl (mM) 2 2 2
MgCl2 (mM) 1 1 1
HEPES (mM) 5 5 5
NMDG-Cl (mM) 87
CaCl2 (mM) 1,8 1,8
Einstellung auf pH 7,4 mit NaOH Tris Tris
Tabelle 2: Lösungen für Biotinylierung und Western Blot
Biotinylierungs
-puffer Glycin-Puffer Lysis-Puffer
Triethanolamin (mM) 10
NaCl (mM) 150 500
CaCl2 (mM) 2
EZ-link sulfo-NHS-SS-
Biotin 1 mg/ml
Glycin (mM) 192
Tris-Cl (mM) 25 50
EDTA (mM) 5
pH 9,5 7,5 7,4

16
Für die Versuche der Kanalaktivierung wurde 2-(Trimethylammonium)-ethyl-
methan-thiosulfonsäure-bromid (MTSET; Toronto Research Chemicals,
Toronto, Kanada), Chymotrypsin (Sigma, Taufkirchen) und S3969 [46]
verwendet. S3969 wurde wie zuvor in der Literatur beschrieben synthetisiert
[32, 46].
4.3 cRNA-Herstellung
Die für die α-, β- und γ-Untereinheit von ENaC kodierende Volllängen-cDNA
war in den pcDNA3.1-Vektor (Invitrogen, Carlsbad, Kalifornien, USA) integriert.
Die Plasmide wurden mit dem Restriktionsenzym Not I (New England Biolabs,
Ipswich, Massachusetts, USA) linearisiert und nach Aufreinigung mit dem
QIAquick PCR Purification Kit (Qiagen, Hilden) als Matrize für die cRNA-
Synthese eingesetzt. Diese wurde mit einem mMESSAGEmMASCHINE Kit T7
(Applied Biosystems, Carlsbad, Kalifornien, USA) durchgeführt, wobei die in
den Vektor integrierte Promotorsequenz des Bakteriophagen T7 als Promotor
für die in vitro Synthese der ENaC-cRNAs diente. Die resultierende cRNA
wurde mit einem NucleoSpin RNA II Kit (Macherey-Nagel, Düren) aufgereinigt
und in nukleasefreiem Wasser aufgenommen. Nach Messung ihrer
Konzentration wurde die fertige cRNA bei -80°C aufbewahrt.
Mittels positionsgerichteter Mutagenese (QuickChange II Site-directed
Mutagenesis Kit, Stratagene, Amsterdam, Niederlande) wurden verschiedene
Mutationen in β-ENaC eingefügt: βS520C, βV348M mit zusätzlicher S520C-
Mutation, βV348M, βV348del, βV348G, βV348L, βV348W, βV348K, βV348D,
βV348E, βV348Q. Die Mutationen wurden per Sequenzanalyse bestätigt.
4.4 Isolation und Injektion der Oozyten
Nachdem die Frösche mit 0,2% MS222 (Sigma) anästhesiert worden waren,
wurden die Oozyten durch partielle Ovariektomie von weiblichen erwachsenen
Xenopus laevis Fröschen gewonnen. Für die Ovariektomie wurde die
Bauchdecke des Frosches durch eine etwa 1 cm lange Inzision eröffnet, die
nach der Entnahme durch Einzelknopfnähte verschlossen wurde. Die Frösche
wurden danach einen Tag in einem separaten Becken außerhalb der
Sammelaquarien gehalten. Bis zur nächsten Oozytenentnahme wurde ein

17
Intervall von mindestens 12 Wochen eingehalten. Die Ovarialläppchen wurden
mechanisch eröffnet und die Oozyten mittels enzymatischem Verdau mit
Kollagenase Typ 2 (CLS2, Biochrom, Berlin, Deutschland; 600-700 U/ml, 3-4h
bei 19°C) aus dem Ovargewebeverband und aus der sie umgebenden
Follikelepithelzellschicht gelöst. Die Kollagenase war hierfür in OR2-Lösung
gelöst. Um die letzten Reste der Follikelepithelzellschicht und die Kollagenase
zu entfernen, wurden die Oozyten nochmals 15 Minuten in Kollagenase-freier
OR2-Lösung geschwenkt.
Oozyten der Stadien V-VI wurden unter dem Stereomikroskop selektiert, wobei
gleichmäßige Pigmentierung und Größe entscheidende Auswahlkriterien
darstellten (Abb. 7). Mit Hilfe eines Mikroinjektors (Nanoject II automatic
injector, Drummond, Broomall, Pennsylvania, USA) wurden die selektierten
Oozyten mit 0,5 ng/ENaC-Untereinheit cRNA injiziert. Hierfür wurden mit Hilfe
eines Mikroelektroden-Pullers (Sutter P-97, Science Products, Hofheim)
Glaskapillaren zu Injektionskapillaren mit langen Spitzen ausgezogen. Diese
wurden unter dem Mikroskop mit einer feinen Pinzette auf einen Durchmesser
von ca. 10-20 μm abgebrochen. Zur gleichmäßigen Druckübertragung durch
den Injektor wurden die Injektionskapillaren mit Mineralöl befüllt. Nach Auftauen
der cRNA-Lösung wurde diese in eine Hämatokritkapillare pipettiert, aus der sie
dann luftblasenfrei in die Injektionskapillare aufgezogen wurde. Die Oozyten
wurden in eine mit ND96 gefüllte Injektionskammer gelegt und die
Injektionskapillare etwa im 45°-Winkel in die Oozyten eingestochen. Pro Oozyte
wurden 46 nl cRNA-Lösung injiziert.
Nach der Injektion wurden die Oozyten in der Regel in einer Lösung mit
niedrigem Na+-Gehalt (ND9) bei 19°C inkubiert. In einigen Versuchen wurden
die Oozyten in einer Lösung mit hohem Na+-Gehalt (ND96) inkubiert. Um
unerwünschtes Bakterienwachstum zu verhindern, wurde den Lösungen 100
U/ml Penicillin und 100 μg/ml Streptomycin zugesetzt.

18
4.5 Elektrophysiologische Methoden
4.5.1 Zwei-Elektroden-Spannungsklemme
Die ENaC-Expression in den Oozyten wurde in der Regel zwei Tage nach
cRNA-Injektion mit der Zwei-Elektroden-Spannungsklemme quantifiziert. Der
schematische Aufbau der Zwei-Elektroden-Spannungsklemme ist in Abb. 8
dargestellt.
Mit einer intrazellulären Elektrode wird das Potenzial der Zelle abgeleitet und
über einen Differenzverstärker mit einem vorgegebenen Haltepotenzial
verglichen. Weicht das Zellpotenzial vom Haltepotenzial ab, wird über eine
zweite intrazelluläre Elektrode ein depolarisierender oder hyperpolarisierender
Ausgleichsstrom appliziert, bis das Zellpotenzial dem Haltepotenzial entspricht.
Über endogene und heterolog exprimierte Ionenkanäle fließen Ladungen ab,
die die Zelle erneut vom Haltepotenzial entfernen. Um dies zu verhindern
müssen folglich kontinuierlich Ausgleichsströme appliziert werden, die diesem
Ladungsfluss entsprechen. Die Messung dieser Ausgleichsströme in An- und
Abwesenheit eines spezifischen Kanalblockers ermöglicht die Quantifizierung
von Strömen durch heterolog exprimierte Ionenkanäle.
In der vorliegenden Arbeit wurde ein OC 725C-Verstärker (Warner Instruments
Corp., Hamden, Conneticut, USA) verwendet, der über ein Interface (LIH 1600,
HEKA, Lambrecht) mit einem Computer verbunden war. Die Ansteuerung des
Verstärkers sowie die Datenerfassung erfolgten per Software (Pulse 8.67,
HEKA). Zusätzlich wurden Strom- und Spannungsdaten auf einem
Zweikanalschreiber (BD12E, Kipp & Zonen, Gengenbach Messtechnik,
Reichenbach/Fils) auf Papier dokumentiert. Die Elektroden bestanden aus mit
einem Mikroelektroden-Puller (Sutter P-97, Science Products, Hofheim)
gezogenen, spitzen Glaskapillaren, die mit 3M KCl-Lösung gefüllt wurden.
Vervollständigt wurde jede Elektrode durch das Eintauchen eines mit dem
Verstärker verbundenen chloridierten Silberdrahtes (Ag/AgCl-Elektrode). Die
nach Eintauchen in die Badlösung gemessenen Elektrodenwiderstände lagen
zwischen 0,3 und 3 MΩ.

19
Abb. 8: Prinzip der Zwei-Elektroden-Spannungsklemme.
4.5.2 Messablauf
Um elektromagnetische Störeinflüsse zu verhindern, wurden die Messungen in
einem von einem Faradayschen Käfig umgebenen Messstand durchgeführt.
Alle Geräte waren geerdet. Die Experimente wurden bei Raumtemperatur
durchgeführt. Die Oozyten wurden in der Mitte einer schmalen Messkammer
positioniert, in der sie kontinuierlich mit Lösung superfundiert wurden. Die
Messlösungen befanden sich in Behältnissen über der Messkammer und
wurden über Silikonschläuche und ein elektromagnetisches Klappensystem
(ALA BPS 8, gesteuert über ein TIB 14 Interface, beide HEKA) zur
Messkammer geleitet. Eine pneumatische Vakuumpumpe (Ejektor SEG,
Schmalz, Glatten) sorgte für kontinuierliches Absaugen der Badlösung.
Unter stereomikroskopischer Kontrolle wurden Strom- und Spannungs-
elektrode über Mikromanipulatoren in die Zelle eingestochen und ein
Haltepotentzial von -60 mV angelegt. Abb. 9A zeigt zwei typische Stromkurven
ENaC exprimierender Oozyten. Die Oozyten wurden konstant mit einer
Flussrate von 2-3 ml/min superfundiert. Die Versuche begannen unter
Superfusion mit 2 μM Amilorid-haltiger ND96-Lösung, um ENaC spezifisch zu
blockieren und somit einen vorzeitigen Na+-Einstrom in die Zellen zu

20
verhindern. Die Amilorid-Superfusion wurde solange durchgeführt, bis sich ein
stabiles Ausgangsniveau des Ganzzellstroms einstellte. Anschließend wurde
Amilorid durch Superfusion mit Amilorid-freier ND96-Lösung aus der
Messkammer ausgewaschen. Nach Auswaschen von Amilorid zeigt sich
zunächst eine einwärts gerichtete Stromkomponente, die von einer teilweisen
Inhibition gefolgt ist, der sogenannten Na+-„self-inhibition“. Dieses Phänomen
hängt von der extrazellulären Na+-Konzentration ab und lässt sich auch durch
rasche Erhöhung der extrazellulären Na+-Konzentration auslösen. Es beruht auf
einer Erniedrigung der Offenwahrscheinlichkeit (Po) von ENaC binnen
Sekunden [14]. Um gegebenenfalls eine unspezifische Veränderung des
Basalstroms („Baselinedrift“) zu erkennen, wurde nach dem Auswaschen von
Amilorid erneut 2 μM Amilorid-haltige ND96-Lösung in die Messkammer
gegeben. Aus der Differenz der Ströme in An- und Abwesenheit von Amilorid
wurde der Amilorid-sensitive Ganzzellstrom ΔIAmi bestimmt (Abb. 9A). ΔIAmi stellt
dabei ein Maß für den durch ENaC vermittelten Einwärtsstrom und damit die
Aktivität von ENaC dar. In nicht injizierten nativen Oozyten ließ sich keine
Amilorid-sensitive Stromkomponente nachweisen.
4.6 Proteinnachweis
4.6.1 Nachweis membranständiger Proteine durch Biotinylierung
Um den an der Zelloberfläche exprimierten Anteil an ENaC isoliert vom
intrazellulär exprimierten Anteil nachzuweisen, können Zellmembranproteine
mit einem zellmembranimpermeablen Biotin-Analogon markiert werden. Diese
Markierung basiert auf kovalenten Bindungsreaktionen zwischen primären
Amingruppen entweder von Lysinresten oder endständiger α-Aminogruppen der
Membranproteine mit den Biotin-Analoga [23]. Biotin besitzt die Eigenschaft
auch hochaffin an Streptavidin und dessen Analoga binden zu können, die sich
wiederum mit sogenannten „Beads“ koppeln lassen. Dabei handelt es sich um
kleine Kügelchen, die den Streptavidin-Biotin-Komplexen eine größere Masse
geben. Dadurch können die Biotin-Streptavidin-Komplexe durch Zentrifugation
von den restlichen, nicht biotinylierten intrazellulären Proteinen abgetrennt
werden und somit ausschließlich biotinylierte Oberflächenproteine separat
nachgewiesen werden.

21
Für die Biotinylierungsversuche in dieser Arbeit wurden jeweils 30 Oozyten pro
untersuchter Gruppe verwendet. Alle Versuchsschritte wurden auf Eis
durchgeführt. Zuerst wurden die Oozyten dreimal in ND96 gewaschen.
Anschließend wurden die Oozyten 15 Minuten lang unter milder Badbewegung
in 1 mg/ml Biotin in Biotinylierungspuffer inkubiert. Um die
Biotinylierungsreaktion zu stoppen und ungebundenes Biotin zu entfernen,
wurden die Oozyten zweimal 5 Minuten lang in Glycin-Puffer bei pH 7,5
gewaschen. Hierauf wurden die Oozyten durch fünfmaliges Aufsaugen durch
eine feine Kanüle (27 Gauge) in Lysis-Puffer lysiert. Zur Vermeidung eines
Proteinabbaus war der Lysis-Puffer mit einer Mischung von Protease-
Inhibitoren („Complete Mini EDTA-free“ protease inhibitor cocktail tablets,
Roche Diagnostics, Mannheim) versetzt. Die Lysate wurden anschließend 10
Minuten bei 1000g zentrifugiert, um Zelldetritus und Zellkerne zu entfernen. Die
Überstände wurden in 1,5 ml-Reaktionsgefäße überführt. Nach Zugabe der
Detergenzien 1% Triton X-100 und 1% Igepal CA-630 (Sigma) wurden die
Überstände für 20 Minuten auf Eis inkubiert und 100 μl Immunopure
immobilized Neutravidin Beads (Pearce, Rockford, Illinois, USA) hinzugefügt,
die vorher durch dreimaliges Aufnehmen in 1 ml Lysis-Puffer und
anschließendem Pelletieren bei 1000g im Lysis-Puffer äquilibriert wurden. Bei
den Neutravidin Beads handelt es sich um ein chemisches Streptavidin-
Analogon, das mit hoher Affinität an Biotin bindet. Die Lysate wurden mit den
Neutravidin Beads über Nacht bei 4°C über Kopf rotierend in Bewegung
gehalten, um die biotinylierten Oberflächenproteine an die Neutravidin Beads zu
koppeln. Nach dieser Inkubation wurden die Reagenzgefäße erneut bei 1000g
3 Minuten lang zentrifugiert, um die mit Biotin-Neutravidin-Komplexen
markierten Oberflächenproteine in Form des Pellets von den nicht-biotinylierten,
intrazellulären Proteinen im Überstand abzutrennen. Das Pellet wurde
anschließend dreimal mit Lysis-Puffer gewaschen.
Zur Kontrolle, ob alle Biotinmoleküle an Neutravidin-Beads gebunden hatten,
wurde aus dem Überstand ein sogenannter „Dot Blot“ angefertigt. Dazu wurde 1
μl des Überstands, in dem sich keine biotinylierten Proteine mehr befinden
sollten, auf eine Nitrozellulose-Membran pipettiert und dort mit
Meerrettichperoxidase gekoppeltem Streptavidin markiert. Die Bindung der
Meerrettichperoxidase wurde dann per Chemilumineszenz nachgewiesen. Die
Experimente wurden nur dann weitergeführt, wenn auf der Membran nahezu

22
kein Signal nachzuweisen war, folglich nahezu alle biotinylierten Proteine an
Neutravidin Beads gebunden hatten und durch Zentrifugation abgetrennt
worden waren. Als Positivkontrolle mit Nachweis eines starken
Chemilumineszenz-Signals diente ein analog durchgeführter „Dot-Blot“ der
biotinylierten Proteine vor Zugabe der Neutravidin Beads.
4.6.2 Proteinnachweis durch Western Blot
Zu den durch die Neutravidin-Beads in Form des Pellets isolierten
Membranproteinen wurden 100 μl eines 2-fach Natriumdodecylsulfat-
Polyacrylamidgelelektrophorese(SDS-PAGE)-Probenpuffers (Rotiload 1, Roth,
Karlsruhe, Deutschland) gegeben. Zur Trennung der Proteine von den Beads
wurden die Proben 5 Minuten bei 95°C erhitzt und 3 Minuten bei 16000g
zentrifugiert. Danach wurde der Überstand auf 8% bzw. 10% SDS-PAGE-Gele
aufgetragen, um die Proteine der Größe nach aufzutrennen. In analoger Weise
wurden auch die aufbewahrten intrazellulären Proteine auf das Gel
aufgetragen, um ebenfalls per Western Blot analysiert zu werden. Zur
Größenanalyse wurde zusätzlich ein „Molekulargewichtsstandard“ (Prestained
Protein Molecular Weight Marker, Fermentas, St. Leon-Rot) aus farblich
markierten Proteinen mit definiertem Molekulargewicht im Gel aufgetrennt. Die
der Größe nach getrennten Proteine wurden hierauf auf eine Polyvinyliden-
Difluorid-Membran durch ein halbtrockenes („semidry“) Elektro-Blotverfahren
transferiert. Dort wurden die Proteine durch eine Antikörper-vermittelte Reaktion
spezifisch nachgewiesen. Für diese Nachweisreaktion wurde die Membran mit
Kaninchen-anti-human-β-ENaC-Antikörper [30, 44] in einer Verdünnung von
1:10000 versetzt. Diese wurden wiederum mittels Meerrettich-Peroxidase-
gekoppelter Ziege-anti-Kaninchen-Antikörper in einer Verdünnung von 1:50000
nachgewiesen (Santa Cruz Biotechnology, Heidelberg), deren Bindung in einer
Chemilumineszenzreaktion sichtbar gemacht werden konnte. Hierfür wurde
„Amersham ECL Plus“ (GE Healthcare, Little Chalfont, UK) verwendet. Ein
Imager (Stella 3200, Raytest, Straubenhardt) detektierte die Intensität der
Chemilumineszenzsignale. Dadurch konnten die nachzuweisenden Proteine
qualitativ und vor allem quantitativ densitometrisch ausgewertet werden.
Zum Nachweis, dass keine intrazellulären Proteine durch Biotin markiert
worden waren, wurden sowohl die aus der Membran stammenden als auch die

23
intrazellulären Proteine mit Antikörpern gegen das ausschließlich intrazellulär
vorkommende Protein β-Aktin in einer Verdünnung von 1:5000 inkubiert
(Sigma). Die Experimente wurden nur dann in die Auswertung eingeschlossen,
wenn in den Western Blots der Membranproteine kein β-Aktin-Signal, bei den
intrazellulären Proteinen dagegen als Positivkontrolle ein starkes β-Aktin-Signal
zu sehen war.
Die quantitative Auswertung der Western Blot Aufnahmen erfolgte mit der
Software ImageJ 1.38x (National Institutes of Health, Bethesda, Maryland,
USA). Dazu wurden die integrierten Intensitätssignale der einzelnen
Proteinbanden auf den unmittelbar benachbarten Hintergrund korrigiert.
4.7 Statistik
Die Daten sind als Mittelwert ± Standardfehler (SEM) dargestellt. Die
statistische Auswertung der Daten erfolgte mit der Software GraphPad Prism
4.03 für Windows (Graph Pad Software, San Diego, Kalifornien, USA). Bei
abhängigen Stichproben wurde der gepaarte t-Test verwendet, bei
unabhängigen Stichproben der ungepaarte t-Test, bei Mehrfachvergleichen
ANOVA mit Dunnett’s Multiple Comparison Test. Die Ergebnisse wurden als
signifikant angenommen, falls die Irrtumswahrscheinlichkeit kleiner 0,05 war.
Die Irrtumswahrscheinlichkeiten wurden wie folgt gekennzeichnet: * p<0,05,
** p<0,01, *** p<0,001. Die Zahlen in oder über den Säulen der Abbildungen
geben die Anzahl n der individuell gemessenen Oozyten an, N die Anzahl
verschiedener Oozyten-Batches, die verwendet wurden.

24
5. Ergebnisse
5.1 Die βV348M-Mutation stimuliert den ENaC-vermittelten Na+-
Strom
Um zu untersuchen, ob die βV348M-Mutante einen stimulatorischen Effekt auf
ENaC ausübt, wurden αβγENaC und αβV348MγENaC in Xenopus laevis Oozyten
exprimiert und mit der Zweielektroden-Spannungsklemme der Amilorid-
sensitive Ganzzellstrom (ΔIami) an Tag 1, 2 und 3 nach der Injektion gemessen.
Abb. 9A zeigt typische Stromkurven einer αβγENaC und einer αβV348MγENaC
exprimierenden Oozyte bei einem Haltepotential von -60 mV. Die Versuche
beginnen in Anwesenheit des spezifischen ENaC-Blockers Amilorid, um eine
vorzeitige Natriumbeladung der Oozyten zu verhindern. Nach Auswaschen von
Amilorid erscheint eine einwärts gerichtete Stromkomponente, die durch den
Einstrom von Na+ hervorgerufen wird. Reapplikation von Amilorid führt den
Strom auf das Ausgangsniveau zurück. Die Differenz der Stromwerte in An- und
Abwesenheit von Amilorid entspricht dem ENaC-vermittelten Na+-Strom ΔIAmi.
In fünf unterschiedlichen Oozyten-Batches ließ sich an αβγENaC
exprimierenden Oozyten einen Tag nach Injektion der cRNA ein ΔIAmi von 0,81
± 0,09 μA (n=60) messen (Abb. 9B). An Tag 2 stieg ΔIAmi 2,7-fach auf 2,19 ±
0,16 μA (n=60), gefolgt von einem weiteren Anstieg an Tag 3 auf 2,66 ± 0,33
μA (3,1-fach im Vergleich zu Tag 1, n=57). αβV348MγENaC exprimierende
Oozyten zeigten an Tag 1 lediglich einen nicht signifikant höheren ΔIAmi (0,99 ±
0,12 µA, n=53) als αβγENaC exprimierende Oozyten. An Tag 2 stieg ΔIami 3,1-
fach auf 3,05 ± 0,29 µA (n=62), gefolgt von einem weiteren Anstieg an Tag 3
auf 3,64 ± 0,47 µA (3,7-fach im Vergleich zu Tag 1, n=57). Somit war an Tag 2
und Tag 3 ein signifikanter stimulatorischer Effekt der βV348M-Mutante von
39,3% bzw. 36,8% im Vergleich zu Wildtyp-ENaC zu verzeichnen.
Da sich der stimulatorische Effekt der βV348M-Mutante an Tag 2 nach der
Injektion als besonders robust erwies, wurden alle weiteren Messungen zu
diesem Zeitpunkt durchgeführt. Um auszuschließen, dass der stimulatorische
Effekt der βV348M-Mutation lediglich auf Unterschieden der cRNA-Qualität
beruhte, wurden die Versuche mit drei verschiedenen cRNA-Batches
durchgeführt. In insgesamt 34 Versuchen mit Oozyten von 34 unterschiedlichen
Fröschen zeigte sich ein signifikanter stimulatorischer Effekt der Mutante auf

25
ΔIAmi von 38% (αβγENaC: 4,03 ± 0,14 µA, αβV348MγENaC: 5,57 ± 0,22 µA, Abb.
9C). Die Ergebnisse demonstrieren somit einen „gain-of-function“-Effekt der
βV348M-Mutante auf die ENaC-Funktion.
10s
wt
1µA
Ami
IAmi
V348M
IAmi
1 2 30
1
2
3
4
5
6
***
wt
V348M
N=5
n=53-62
Tag nach Injektion
I A
mi (
no
rma
lisie
rt)
wt V348M0
1
2
3
4
5
6 ***N=34
331 332
I A
mi (
µA
)
A
B C
Abb. 9: Die βV348M-Mutation stimuliert den ENaC-vermittelten Na+-Strom (mod. nach
[55]). Oozyten wurden mit cRNA für αβγENaC (wt) oder αβV348MγENaC (V348M) injiziert (0,5
ng/Untereinheit) und der Amilorid-sensitive Strom (ΔIAmi) mit der Zwei-Elektroden-
Spannungsklemme detektiert. A, Repräsentative Ganzzell-Stromkurven einer αβγENaC
exprimierenden Oozyte (links) bzw. einer αβV348MγENaC exprimierenden Oozyte (rechts) bei
einem Haltepotential von -60mV. Die Anwesenheit von Amilorid (2 μM) in der Badlösung ist
durch schwarze Balken gekennzeichnet. B, ΔIAmi von αβγENaC und αβV348MγENaC
exprimierenden Oozyten an Tag 1, 2 und 3 nach Injektion. Jeder Datenpunkt repräsentiert den
Mittelwert aus 53-62 individuellen Oozyten aus fünf unterschiedlichen Oozyten-Batches. Die
Messwerte der individuellen Experimente wurden auf den Mittelwert der jeweiligen Wildtyp-
Kontrolle an Tag 1 normalisiert. C, Zusammenfassung aller Experimente an Tag 2 nach
Injektion. Ungepaarter t-Test.

26
5.2 Der stimulatorische Effekt der βV348M-Mutation ist vom
Na+-Gehalt der Oozyten weitgehend unabhängig
Von einer durch Kellenberger et al. untersuchten ENaC-Mutante, die bei
betroffenen Patienten ein Liddle Syndrom verursacht, ist bekannt, dass sich der
stimulatorische Effekt der „gain-of-function“-Mutante im Vergleich zum Wildtyp
erst nach Inkubation der Oozyten in einer Lösung mit hohem Na+-Gehalt und
dadurch gesteigerter intrazellulärer Na+-Konzentration zeigt [36]. Daher sollte
untersucht werden, ob der stimulatorische Effekt der βV348M-Mutation durch
Inkubation der Oozyten in einer Lösung mit hohem Na+-Gehalt (ND96)
vergrößert wird.
Auch in ND96 inkubierte Oozyten zeigten einen messbaren ΔIAmi (Abb. 10). Im
Vergleich zu Oozyten, die in einer Lösung mit niedrigem Na+-Gehalt (ND9)
inkubiert wurden, war ΔIAmi geringer. ΔIAmi des Wildtyps belief sich dabei auf
1,39 ± 0,19 μA, während die Mutante einen etwa 54% größeren ΔIAmi von 2,14
± 0,27 μA aufwies. Der stimulatorische Effekt der Mutante war nach Inkubation
mit ND96 etwas größer als nach Inkubation mit ND9. Im Vergleich zur Liddle-
Mutante nach Inkubation mit einer Lösung mit hohem Na+-Gehalt, die unter
diesen Bedingungen eine etwa 3-fache Steigerung von ΔIAmi im Vergleich zu
Wildtyp-ENaC zeigte [36], war der stimulatorische Effekt der βV348M-Mutation
allerdings deutlich geringer.
Die Auswirkung der intrazellulären Na+-Konzentration auf den stimulatorischen
Effekt der βV348M-Mutante ist also viel weniger ausgeprägt als bei der Liddle-
Mutation und wurde im Folgenden daher nicht weiter untersucht.
Abb. 10: Der stimulatorische Effekt der βV348M-Mutation ist vom Na+-Gehalt der Oozyten
weitgehend unabhängig. ΔIAmi von αβγENaC (wt) und αβV348MγENaC (V348M) exprimierenden
Oozyten wurde nach Inkubation in ND96 2 Tage nach cRNA-Injektion (0,5 ng/Untereinheit)
gemessen. Ungepaarter t-Test.
wt V348M0.0
0.5
1.0
1.5
2.0
2.5 *N=6
58 58
I A
mi (
µA
)

27
5.3 Die βV348M-Mutation erhöht die mittlere Po von ENaC
Die Effekte von „gain-of-function“-Mutationen können prinzipiell auf drei
unterschiedlichen Mechanismen beruhen. Dabei können die Anzahl der in der
Membran exprimierten Kanäle, die Einzelkanalleitfähigkeit oder die
Offenwahrscheinlichkeit Po erhöht sein. Veränderungen der
Einzelkanalleitfähigkeit von humanem ENaC durch Mutationen wurden
allerdings bisher nicht beschrieben.
Zur Bestimmung der mittleren Po wurde in der β-Untereinheit von Wildtyp-ENaC
und auch der βV348M-Mutante an Position 520 das an dieser Stelle
vorkommende Serin durch Cystein substituiert. Bindung des Sulfhydryl-
Reagenz MTSET an dieses nahe der Porenregion von ENaC gelegene Cystein
erhöht die Po auf nahe 1. Dies geschieht durch das Einfügen einer positiven
Ladung und eine daraus resultierende Destabilisierung des Geschlossen-
Zustands des Kanals. [37, 66].
Abb. 11A zeigt repräsentative Stromkurven der Aktivierung mit MTSET (1mM)
sowohl einer αβS520CγENaC als auch einer αβV348M,S520CγENaC exprimierenden
Oozyte. Nach Auswaschen von Amilorid zeigt sich eine einwärts gerichtete
Stromkomponente, die durch Applikation von MTSET noch vergrößert wird. Der
Einwärtsstrom erreicht nach etwa fünf Minuten ein Plateau.
In αβS520CγENaC exprimierenden Oozyten zeigte sich im Mittel aller
untersuchten Oozyten ein ΔIAmi von 4,12 ± 0,33 µA (Abb. 11B). Applikation von
MTSET erhöhte ΔIAmi auf 16,29 ± 0,86 µA, was einer 4,0-fachen Steigerung
entspricht (Abb. 11BC). Unter der Annahme, dass MTSET die Po aller Kanäle
auf 1 setzt, ergibt sich somit für αβγENaC eine mittlere Po von 0,25 vor
Applikation von MTSET.
αβV348M,S520CγENaC exprimierende Oozyten wiesen einen ΔIAmi von 5,88 ± 0,47
μA auf, was einer Stimulation von 43% gegenüber dem ΔIAmi des Wildtyps
entspricht und den stimulatorischen Effekt der Mutante in diesen Experimenten
bestätigt. MTSET erhöhte ΔIAmi auf 16,40 ± 1,03 µA und war somit 2,8-fach
gesteigert (Abb. 11BC). Unter der Annahme, dass MTSET die Po aller Kanäle
auf 1 setzt, ergibt sich somit eine mittlere Po der Mutante von 0,36. Die durch
die βV348M-Mutation verursachte Erhöhung der Po belief sich folglich auf 44%
und entspricht somit dem in diesen Experimenten gefundenen stimulatorischen
Effekt auf ΔIAmi.

28
Interessanterweise liegt ΔIAmi von αβS520CγENaC und αβV348M,S520CγENaC
exprimierenden Oozyten nach MTSET-Stimulation auf gleichem Niveau (Abb.
11B). Dies deutet darauf hin, dass sich die Anzahl der Kanäle in der Membran
zwischen Wildtyp und Mutanten nicht unterscheidet.
Zusammenfassend zeigen diese Ergebnisse, dass eine Erhöhung der Po
entscheidend zum stimulatorischen Effekt der βV348M-Mutante beiträgt.

29
wt
2 µA
60s
Ami AmiMTSET
V348MAmi AmiMTSET
0
5
10
15
20
25
wt V348M
- +MTSET
- +
******
I A
mi (
µA
)
wt V348M0
1
2
3
4
N=3
23 23
***
I A
mi+
MT
SE
T/
I Am
i-in
itia
l
B C
A
Abb. 11: Die βV348M-Mutation erhöht die mittlere Po von ENaC (mod. nach [55]). Oozyten
wurden mit cRNA für αβS520CγENaC (wt) oder αβV348M,S520CγENaC (V348M) injiziert (0,5
ng/Untereinheit). Zur maximalen Erhöhung der Po wurden die Kanäle mit MTSET (1 mM)
aktiviert. A, Repräsentative Ganzzell-Stromkurven von αβS520CγENaC (links) bzw.
αβV348M,S520CγENaC (rechts) exprimierenden Oozyten. Die Anwesenheit von Amilorid (2 µM,
schwarzer Balken) bzw. MTSET (weißer Balken) ist entsprechend gekennzeichnet. B,
Zusammenfassung von zu A analog durchgeführten Experimenten aus drei unterschiedlichen
Oozyten-Batches. Zwei durch eine Linie verbundene Datenpunkte repräsentieren eine
individuell gemessene Oozyte, deren ΔΙAmi vor MTSET-Stimulation durch eine Linie mit dem
zugehörigen ΔΙAmi nach MTSET-Stimulation verbunden ist. Die weiße Säule repräsentiert den
Mittelwert von ΔΙAmi aller Oozyten vor MTSET-Stimulation, die schwarze Säule den Mittelwert
von ΔΙAmi nach MTSET-Stimulation. C, Relative Aktivierung von ΔIAmi durch MTSET dargestellt
als Quotient aus ΔIAmi nach (ΔIAmi+MTSET) und vor (ΔIAmi-initial) MTSET-Aktivierung. Gepaarter
(B) und ungepaarter (C) t-Test.

30
5.4 Chymotrypsin stimuliert αβγENaC mehr als αβV348MγENaC
Die proteolytische Aktivierung von ENaC spielt eine bedeutende Rolle in der
Kanalregulation von ENaC [60]. Daher sollte überprüft werden, ob die βV348M-
Mutation die Aktivierung von ENaC durch Proteasen beeinflusst. Zu diesem
Zweck wurden Versuche durchgeführt, in denen der Kanal mit Hilfe der
Serinprotease Chymotrypsin aktiviert wurde. Chymotrypsin aktiviert ENaC dabei
durch einen bisher nicht völlig aufgeklärten Mechanismus.
Der Versuchsablauf erfolgte analog zur Stimulation mit MTSET. Dabei wurde
die maximale Stimulation durch Chymotrypsin (2 μg/ml) nach etwa fünf Minuten
erreicht.
Abb. 12A zeigt repräsentative Stromkurven der Aktivierung mit Chymotrypsin
sowohl einer αβγENaC als auch einer αβV348MγENaC exprimierenden Oozyte.
Nach Auswaschen von Amilorid zeigt sich eine einwärts gerichtete
Stromkomponente, die durch Applikation von Chymotrypsin noch vergrößert
wird.
In αβγENaC exprimierenden Oozyten zeigte sich im Mittel aller untersuchten
Oozyten ein ΔIAmi von 3,46 ± 0,24 µA. Applikation von Chymotrypsin
vergrößerte ΔIAmi auf 13,40 ± 0,85 µA, was einer 3,9-fachen Steigerung
entspricht (Abb. 12BC). Dieser Effekt entspricht der Aktivierung durch MTSET
und zeigt, dass Chymotrypsin den Kanal ebenfalls vollständig aktiviert.
αβV348MγENaC exprimierende Oozyten wiesen einen ΔIAmi von 4,71 ± 0,33 μA
vor Chymotrypsin-Applikation auf, was einer Stimulation von 36% gegenüber
dem ΔIAmi des Wildtyps entspricht. Dies bestätigt den stimulatorischen Effekt
der Mutante in diesen Experimenten. Chymotrypsin erhöhte ΔIAmi auf 13,36 ±
0,85 µA nach Applikation von Chymotrypsin. Dies entspricht einer 2,8-fachen
Steigerung der Ströme und ist signifikant geringer als die Aktivierung des
Wildtyps durch Chymotrypsin (Abb.12 BC). Die Aktivierung der Mutante durch
Chymotrypsin liegt dabei im Bereich der Aktivierung durch MTSET und zeigt,
dass auch der mutierte Kanal in vollem Umfang durch Chymotrypsin aktivierbar
ist.
Interessanterweise gleichen sich ΔIAmi von αβγENaC und αβV348MγENaC
exprimierenden Oozyten nach Aktivierung durch Chymotrypsin (Abb. 12B). Dies
deutet, wie in den Versuchen mit MTSET, darauf hin, dass sich die Anzahl der
Kanäle in der Zellmembran zwischen Wildtyp und Mutante nicht unterscheidet.

31
Zusammenfassend ist festzuhalten, dass sowohl Wildtyp- als auch βV348M-
ENaC durch Chymotrypsin aktiviert werden kann, der mutierte Kanal jedoch
signifikant geringer. Auch dieser Befund spricht dafür, dass die Mutation eine
Erhöhung der Po bewirkt.
wt
2 µA
60s
Ami AmiChymotrypsin
V348M
Ami AmiChymotrypsin
0
10
20
30
40
wt V348M
- +Chymotrypsin
- +
*** ***
I A
mi (
µA
)
wt V348M0
1
2
3
4
N=9
71 71
***
I Am
i+ch
ym
o/
I Am
i-in
itia
l
B C
A
Abb. 12: Chymotrypsin stimuliert αβγENaC mehr als αβV348MγENaC (mod. nach [55]).
Oozyten wurden mit cRNA für αβγENaC (wt) oder αβV348MγENaC (V348M) injiziert (0,5
ng/Untereinheit). Zur proteolytischen Aktivierung wurden die Kanäle mit Chymotrypsin (2 μg/ml)
stimuliert. A, Repräsentative Ganzzell-Stromkurven von αβγENaC (links) bzw. αβV348MγENaC
(rechts) exprimierenden Oozyten. Die Anwesenheit von Amilorid (2 µM, schwarzer Balken) bzw.
Chymotrypsin (weißer Balken) ist entsprechend gekennzeichnet. B, Zusammenfassung von zu
A analog durchgeführten Experimenten aus neun unterschiedlichen Oozyten-Batches. Zwei
durch eine Linie verbundene Datenpunkte repräsentieren eine individuell gemessene Oozyte,
deren ΔΙAmi vor Chymotrypsin-Stimulation durch eine Linie mit dem zugehörigen ΔΙAmi nach
Chymotrypsin-Stimulation verbunden ist. Die weiße Säule repräsentiert den Mittelwert von ΔΙAmi
aller Oozyten vor Chymotrypsin-Stimulation, die schwarze Säule den Mittelwert von ΔΙAmi nach
Chymotrypsin-Stimulation. C, Relative Aktivierung von ΔΙAmi durch Chymotrypsin dargestellt als
Quotient aus ΔΙAmi nach (ΔΙAmi+chymo) und vor (ΔΙAmi-initial) Chymotrypsin-Aktivierung.
Gepaarter (B) und ungepaarter (C) t-Test.

32
5.5 Die βV348M-Mutante wird durch S3969 weniger aktiviert als
der Wildtyp
Kürzlich wurde von Lu et al. [46] der neue ENaC-Aktivator S3969 vorgestellt.
Interessanterweise betonen Lu et al. die funktionelle Bedeutung der
Aminosäure βV348 für die Aktivierbarkeit durch den Aktivator. Nach Deletion
dieser Aminosäure ließ sich der Kanal nicht mehr durch den Aktivator
stimulieren. Daher sollte untersucht werden, ob die βV348M-Mutation den
stimulatorischen Effekt von S3969 reduziert.
Der Versuchsablauf erfolgte analog zur Stimulation mit MTSET bzw.
Chymotrypsin. Dabei wurde die maximale Stimulation durch S3969 (10 μM)
nach etwa drei bis vier Minuten erreicht.
Abb. 13A zeigt repräsentative Stromkurven einer αβγENaC und einer
αβV348MγENaC exprimierenden Oozyte. Nach Auswaschen von Amilorid zeigt
sich eine einwärts gerichtete Stromkomponente, die durch Applikation von
S3969 noch vergrößert wird.
In allen untersuchten αβγENaC exprimierenden Oozyten zeigte sich im Mittel
ein ΔIAmi von 6,81 ± 0,54 µA. Applikation von S3969 vergrößerte ΔIAmi auf 22,45
± 1,70 µA, was einer 3,3-fachen Steigerung entspricht (Abb. 13BC).
αβV348MγENaC exprimierende Oozyten wiesen einen ΔIAmi von 8,76 ± 1,03 μA
vor S3969-Applikation auf, was einer Stimulation von 29% gegenüber dem ΔIAmi
des Wildtyps entspricht. Dies bestätigt den stimulatorischen Effekt der Mutante
in diesen Experimenten. S3969 erhöhte ΔIAmi auf 20,91 ± 2,18 µA. Dies
entspricht einer 2,4-fachen Steigerung der Ströme und ist signifikant geringer
als die Aktivierung des Wildtyps durch S3969 (Abb. 13BC).
Interessanterweise gleichen sich ΔIAmi αβγENaC und αβV348MγENaC
exprimierender Oozyten nach Aktivierung durch S3969 (Abb. 13B). Dies deutet
erneut darauf hin, dass sich die Anzahl der Kanäle in der Membran zwischen
Wildtyp und Mutante nicht unterscheidet.
Zusammenfassend ist festzuhalten, dass S3969 sowohl αβγENaC als auch
αβV348MγENaC aktiviert. Der etwas stärkere Effekt auf den Wildtyp-Kanal spricht
wiederum für eine erhöhte Po des mutierten Kanals. Das Ausmaß der
Aktivierung durch S3969 war aber insgesamt geringer als durch MTSET- und
Chymotrypsin-Applikation.

33
wt
5 µA
60s
Ami AmiS3969
V348MAmi AmiS3969
0
10
20
30
40
50
wt V348M
- +S3969
- +
******
I A
mi (
µA
)
wt V348M0
1
2
3
4N=5
40 40
***
I A
mi+
S3
96
9/
I Am
i-in
itia
l
B C
A
Abb. 13: Die βV348M-Mutante wird durch S3969 weniger aktiviert als der Wildtyp (mod.
nach [55]). Oozyten wurden mit cRNA für αβγENaC (wt) oder αβV348MγENaC (V348M) injiziert
(0,5 ng/Untereinheit). Die Kanäle wurden durch S3969 (10 μM) aktiviert. A, Repräsentative
Ganzzell-Stromkurven von αβγENaC (links) bzw. αβV348MγENaC (rechts) exprimierenden
Oozyten. Die Anwesenheit von Amilorid (2 µM, schwarzer Balken) bzw. S3969 (weißer Balken)
ist entsprechend gekennzeichnet. B, Zusammenfassung von zu A analog durchgeführten
Experimenten aus fünf unterschiedlichen Oozyten-Batches. Zwei durch eine Linie verbundene
Datenpunkte repräsentieren eine individuell gemessene Oozyte, deren ΔΙAmi vor S3969-
Stimulation durch eine Linie mit dem zugehörigen ΔΙAmi nach S3969-Stimulation verbunden ist.
Die weiße Säule repräsentiert den Mittelwert von ΔΙAmi vor S3969-Stimulation, die schwarze
Säule den Mittelwert von ΔΙAmi aller Oozyten nach S3969-Stimulation. C, Relative Aktivierung
von ΔIAmi durch S3969 dargestellt als Quotient aus ΔΙAmi nach (ΔΙAmi+S3969) und vor (ΔΙAmi-
initial) S3969-Aktivierung. Gepaarter (B) und ungepaarter (C) t-Test.

34
Aus diesem Grund sollte geklärt werden, ob S3969 im Gegensatz zu
Chymotrypsin den Kanal nicht vollständig aktiviert. Zu diesem Zweck wurden
αβγENaC exprimierende Oozyten mit Chymotrypsin (2μg/ml) und S3969 (10
μM) sequentiell stimuliert.
Abb. 14A zeigt zwei repräsentative Stromkurven aus diesen Experimenten. In
der linken Stromkurve wurde nach Auswaschen von Amilorid für etwa fünf
Minuten Chymotrypsin appliziert, bis der Einwärtsstrom ein Plateau ausbildete.
Unmittelbar folgend wurde zusätzlich S3969 in die Superfusionslösung
gegeben, um eine weitere Aktivierung durch S3969 feststellen zu können.
Hierbei ließ sich kein zusätzlicher Anstieg der Ströme durch S3969 beobachten.
An insgesamt 5 Oozyten ergab sich als Mittelwert für ΔIAmi vor Stimulation 10,28
± 1,38 μA, für ΔIAmi nach Chymotrypsin-Stimulation 27,22 ± 3,59 μA und für
ΔIami nach zusätzlicher Stimulation mit S3969 26,70 ± 3,66 μA.
Als Kontrollexperiment wurde die Reihenfolge der Zugabe von Chymotrypsin
und S3969 getauscht. Nach Auswaschen von Amilorid ließ sich ein ΔIAmi von
5,34 ± 0,87 μA messen. Anschließend erfolgte eine drei- bis vierminütige
S3969-Superfusion, die ΔIAmi auf 17,77 ± 1,55 μA steigerte. Zusätzliche
Stimulation durch Chymotrypsin erbrachte einen weiteren Anstieg von ΔIAmi um
8% auf 19,16 ± 1,99 μA (p= 0,11; Abb. 14A, rechte Stromkurve). Abb. 14B zeigt
die Zusammenfassung der Stromwerte aus beiden Versuchsanordnungen.
Die Ergebnisse zeigen einen nicht signifikanten Trend, dass S3969 im
Vergleich zu Chymotrypsin den Kanal nicht vollständig aktiviert. Dieser Trend
war bei jeder untersuchten Oozyte zu finden.

35
Abb. 14: Chymotrypsin aktiviert Wildtyp-ENaC stärker als S3969. Oozyten wurden mit
cRNA für αβγENaC injiziert (0,5 ng/Untereinheit). Zur Aktivierung der Kanäle wurde S3969 (10
μM) bzw. Chymotrypsin (2 μg/ml) alleine oder zusammen appliziert. A, Repräsentative
Ganzzell-Stromkurven von αβγENaC exprimierenden Oozyten. Die linke Stromkurve zeigt nach
Auswaschen von Amilorid (2 µM) die Aktivierung mit Chymotrypsin, der eine zusätzliche
Superfusion mit S3969 folgt. Die rechte Stromkurve zeigt den umgekehrten Fall: Wildtyp-ENaC
wird nach Auswaschen von Amilorid mit S3969 und hierauf zusätzlich mit Chymotrypsin
aktiviert. Die Anwesenheit von Amilorid und Chymotrypsin alleine oder zusammen ist
entsprechend gekennzeichnet. B, Zusammenfassung aller zu A analog durchgeführten
Messungen. Linkes Diagramm: Analog zur linken Stromkurve durchgeführte Experimente.
Rechtes Diagramm: Analog zur rechten Stromkurve durchgeführte Experimente. Jeder
Datenpunkt repräsentiert eine individuell gemessene Oozyte, deren ΔΙAmi vor Stimulation durch
eine Linie mit dem zugehörigen ΔΙAmi nach S3969- bzw. Chymotrypsin-Stimulation verbunden
ist. ΔΙAmi nach S3969- bzw. Chymotrypsin-Stimulation ist hierauf mit dem zugehörigen ΔΙAmi
nach Stimulation mit S3969 und gleichzeitig Chymotrypsin verbunden. Gepaarter t-Test.
Ami
IAmi
IAmi+Chymo IAmi+Chymo/S3969
Chymo Chymo/S3969
5 µA
60s
Ami
IAmi
IAmi+S3969 IAmi+S3969/Chymo
S3969 S3969/Chymo
Kontrolle
+ Chym
o
+ Chym
o/S3969
0
10
20
30
40
50 n=5
I A
mi (
A)
Kontrolle
+ S3969
+ S3969/Chym
o
0
5
10
15
20
25
n=5
I A
mi (
A)
A
B
Ami
IAmi
IAmi+Chymo IAmi+Chymo/S3969
Chymo Chymo/S3969
5 µA
60s
Ami
IAmi
IAmi+S3969 IAmi+S3969/Chymo
S3969 S3969/Chymo
Kontrolle
+ Chym
o
+ Chym
o/S3969
0
10
20
30
40
50 n=5
I A
mi (
A)
Kontrolle
+ S3969
+ S3969/Chym
o
0
5
10
15
20
25
n=5
I A
mi (
A)
A
B
A
B

36
5.7 Verschiedene Mutationen an βV348
5.7.1 Aminosäure-Mutationen zeigen unterschiedliche Effekte auf die
ENaC-Funktion
Die Methionin-Seitenkette an Position 348 der β-Untereinheit des mutierten
Kanals besitzt im Vergleich zu der an dieser Stelle im Wildtyp-Kanal
vorkommenden Valin-Seitenkette aufgrund der zusätzlichen -S-CH3-Gruppe
andere Größenverhältnisse. Daher wurden Versuche durchgeführt, um
herauszufinden, ob die Größe oder die Ladung der Aminosäureseitenkette an
Position 348 der β-Untereinheit systematisch die ENaC-Funktion verändert.
Hierfür wurden mit positionsgerichteter Mutagenese verschiedene Mutanten
generiert, die sich sowohl in Ladung als auch Größe der Aminosäuren-
Seitenkette (Abb. 15) unterschieden.
Dabei wurde Valin durch die viel kleinere Aminosäure Glycin (V348G), die
verzweigtkettige Aminosäure Leucin (V348L) oder die aromatische Aminosäure
Tryptophan (V348W) ersetzt. Zudem wurden sowohl die sich in ihrer Länge um
eine CH3-Gruppe unterscheidenden negativ geladenen Aminosäuren Aspartat
(V348D) und Glutamat (V348E), als auch die positiv geladene Aminosäure
Lysin (V348K) als Ersatz für das Valin verwendet. Als weitere Mutante wurde
das Amid Glutamin untersucht (V348Q). Außerdem wurde V348 deletiert
(V348del).
Abb. 15: Aminosäuren, die durch positionsgerichtete Mutagenese zur Generierung
verschiedener Mutanten eingesetzt wurden.
O
NH2 OH
O
NH2
NH2 OH
O
NH2
SOH
O
NH2
OH
O
NH2NH
OH
O O
OHOH
NH2
O
NH2
OH
OHOH
NH2O
O
O
O
NH2
NH2
OH
Lysin (K) Glycin (G)
O
NH2 OH
O
NH2
NH2 OH
O
NH2
SOH
O
NH2
OH
O
NH2NH
OH
O O
OHOH
NH2
O
NH2
OH
OHOH
NH2O
O
O
O
NH2
NH2
OH
Leucin (L)
O
NH2 OH
O
NH2
NH2 OH
O
NH2
SOH
O
NH2
OH
O
NH2NH
OH
O O
OHOH
NH2
O
NH2
OH
OHOH
NH2O
O
O
O
NH2
NH2
OH
Tryptophan (T)
O
NH2 OH
O
NH2
NH2 OH
O
NH2
SOH
O
NH2
OH
O
NH2NH
OH
O O
OHOH
NH2
O
NH2
OH
OHOH
NH2O
O
O
O
NH2
NH2
OH
Glutamat (E)
O
NH2 OH
O
NH2
NH2 OH
O
NH2
SOH
O
NH2
OH
O
NH2NH
OH
O O
OHOH
NH2
O
NH2
OH
OHOH
NH2O
O
O
O
NH2
NH2
OH
Glutamin (Q)
O
NH2 OH
O
NH2
NH2 OH
O
NH2
SOH
O
NH2
OH
O
NH2NH
OH
O O
OHOH
NH2
O
NH2
OH
OHOH
NH2O
O
O
O
NH2
NH2
OH
Aspartat (D)
O
NH2 OH
O
NH2
NH2 OH
O
NH2
SOH
O
NH2
OH
O
NH2NH
OH
O O
OHOH
NH2
O
NH2
OH
OHOH
NH2O
O
O
O
NH2
NH2
OH
Valin (V)
O
NH2 OH
O
NH2
NH2 OH
O
NH2
SOH
O
NH2
OH
O
NH2NH
OH
O O
OHOH
NH2
O
NH2
OH
OHOH
NH2O
O
O
O
NH2
NH2
OH
Methionin (M)
O
NH2 OH
O
NH2
NH2 OH
O
NH2
SOH
O
NH2
OH
O
NH2NH
OH
O O
OHOH
NH2
O
NH2
OH
OHOH
NH2O
O
O
O
NH2
NH2
OH

37
Für die Untersuchung der Effekte auf ΔIAmi der einzelnen Mutanten wurden
jeweils zwei unterschiedliche cRNA-Batches verwendet, die vergleichbare
Ergebnisse lieferten. Abb. 16 zeigt die Zusammenfassung dieser Versuche. Die
verschiedenen Mutanten sind dabei nach der Größe des Effekts auf ΔIAmi
angeordnet.
Dabei stimulierte die kleinste und ungeladene Aminosäure Glycin genauso wie
die größere und positiv geladene Aminosäure Lysin ΔIAmi stärker als die
βV348M-Mutation. Einen ebenfalls stimulatorischen, aber geringeren Effekt als
die βV348M-Mutation übten das ungeladene Leucin und das aromatische
Tryptophan auf ΔIAmi aus.
Das negativ geladene Glutamat hatte dagegen keinen Effekt auf die ENaC-
Funktion im Vergleich zum Wildtyp. Das ebenfalls negativ geladene Aspartat
zeigte sogar einen geringen inhibitorischen Effekt.
Interessanterweise bewirkte die Insertion der polaren Aminosäure Glutamin in
ähnlicher Weise wie die Deletion des Valins an Position 348 der β-Untereinheit
einen beinahe vollständigen Verlust der ENaC-Funktion.
Insgesamt ergab sich in den Versuchen der an Position 348 der β-Untereinheit
mutierten Kanäle kein systematischer Zusammenhang zwischen dem Effekt auf
ΔIAmi und Ladung oder Größe der Aminosäure.
Abb. 16: Aminosäure-Mutationen zeigen unterschiedliche Effekte auf die ENaC-Funktion
(mod. nach [55]). Die Oozyten wurden mit cRNA von Wildtyp-ENaC (wt) oder der
unterschiedlichen Mutanten (wie unter den Säulen angegeben) injiziert (0,5 ng/Untereinheit).
ΔIAmi wurde 2 Tage nach Injektion bestimmt. Um die Daten aus verschiedenen Batches Oozyten
zusammenzufassen, wurde ΔIAmi der Mutanten auf den durchschnittlichen ΔIAmi der
entsprechenden Wildtyp-ENaC (wt) exprimierenden Oozyten normalisiert. ANOVA mit Dunnett’s
Multiple Comparison Test gegen Wildtyp-Kontrolle.
V34
8G
V34
8K
V34
8M
V34
8L
V34
8W
V34
8E wt
V34
8D
V34
8Q
V34
8del
0.0
0.5
1.0
1.5
2.0
249178 749191 120121 92121 90
N=9-28***
******
*** ***
*
*** *** I
Am
i(n
orm
alis
iert
)

38
5.7.2 Effekte der Mutationen auf die Proteinexpression
Um zu untersuchen, ob die Effekte der Mutanten auf ΔIAmi auf einer veränderten
Proteinexpression des Kanals intrazellulär oder in der Membran beruht, wurde
in weiteren Versuchen parallel zur Bestimmung von ΔIAmi in Oozyten aus der
gleichen Präparation die Proteinexpression von β-ENaC bestimmt.
Die intrazelluläre Proteinexpression zeigte mit Ausnahme von V348Q und
V348del keine signifikanten Unterschiede zwischen Wildtyp und den
unterschiedlichen Mutanten (Abb. 17B). V348Q und V348del wurden signifikant
geringer exprimiert als der Wildtyp.
Die Proteinexpression in der Membran zeigte mit Ausnahme von V348Q und
V348del ebenfalls keine signifikanten Unterschiede zwischen Wildtyp und den
unterschiedlichen Mutanten (Abb. 17C). V348Q und V348del wurden auch in
der Membran signifikant geringer exprimiert als der Wildtyp.
Die Effekte der Mutationen auf ΔIAmi lassen sich somit nicht durch Unterschiede
in der Proteinexpression intrazellulär oder in der Membran erklären, mit
Ausnahme von V348Q und V348del. Eine Erhöhung der in die Membran
integrierten ENaC-Moleküle scheint somit auch nicht zum „gain-of-function“-
Effekt der βV348M-Mutante beizutragen.

39
V34
8G
V34
8K
V34
8M
V34
8L
V34
8W
V34
8E wt
V34
8D
V34
8Q
V34
8del
0.0
0.5
1.0
1.5
2.0
**
4 48 44 48 444
*****
E
NaC
intrazellu
lär (n
orm
.)
V348G
V348K
V348M
V348L
V348W
V348E w
t
V348D
V348Q
V348d
el
0.0
0.5
1.0
1.5
2.0
*** 4 48 44 48 444** **
E
NaC
in M
em
bra
n (norm
.)
V34
8G
V34
8K
V34
8M
V34
8L
V34
8W
V34
8E wt
V34
8D
V34
8Q
V34
8del
0.0
0.5
1.0
1.5
2.0
2.5
8282 404040 4042 4242 42
N=4-8***
****** *****
******
I
Am
i(n
orm
.)
A
B
C
D

40
Abb. 17: Auswirkungen der unterschiedlichen Mutationen auf die βENaC-Expression
intrazellulär und in der Zellmembran (mod. nach [55]). Die Oozyten wurden mit cRNA von
Wildtyp-ENaC (wt) oder der unterschiedlichen Mutanten (wie angegeben) injiziert (0,5
ng/Untereinheit) und für zwei Tage inkubiert. Intrazelluläres und aus der Zellmembran
stammendes Protein wurden mit SDS-PAGE aufgetrennt und β-ENaC durch spezifische
Antikörper nachgewiesen. A, Repräsentative Western Blots. B und C, Experimente wie in A
wurden viermal durchgeführt und die Signale mittels densitometrischer Analyse quantifiziert. Die
Zahlen in den Säulen geben hier die Anzahl an untersuchter Oozyten-Batches an. D, ΔIAmi der
unter den Säulen gekennzeichneten βV348-Mutanten aus den Experimenten von B und C. Um
die Variabilität von Batch zu Batch zu korrigieren, wurden die Daten jeweils auf die
entsprechende Wildtyp-Kontrolle normalisiert. ANOVA mit Dunnett’s Multiple Comparison Test
gegen Wildtyp-Kontrolle.

41
5.7.3 Proteolytische Aktivierung der mutierten Kanäle mit Chymotrypsin
Da sich die verschiedenen Mutanten, mit Ausnahme der beiden stark
inhibitorischen Mutanten V348Q und V348del, in der Proteinexpression von β-
ENaC in der Membran nicht unterschieden, führen wahrscheinlich Unterschiede
der mittleren Po zu den veränderten ΔIAmi-Werten. Um dies nachzuweisen,
wurden die einzelnen Mutanten mit Ausnahme von V348Q und V348del (wegen
mangelnder Expression in der Zellmembran) mit Chymotryspin stimuliert. Wie
gezeigt (vgl. 5.4) aktiviert Chymotrypsin ΔIAmi maximal. Unterschiede der Po der
Mutanten können demnach durch Unterschiede in der relativen Aktivierung
durch Chymotrypsin erfasst werden. Der Versuchsablauf und die Auswertung
der Daten erfolgte wie in 5.4 beschrieben.
Die Ergebnisse sind in Abb. 18A zusammengefasst. ΔIAmi erreichte nach der
Aktivierung mit Chymotrypsin bei allen Mutanten ähnliche Werte, was die
ähnliche Expression der Mutanten in der Oberflächenmembran anzeigt. In Abb.
18B sind die relativen stimulatorischen Effekte gegen die normalisierten
„Baseline“-ΔIAmi-Werte vor Aktivierung mit Chymotrypsin aufgetragen. Die
Abbildung zeigt, dass ein höherer „Baseline“-ΔIAmi mit einer geringeren relativen
Aktivierung durch Chymotrypsin einhergeht. Dies deutet darauf hin, dass die bei
einigen Mutanten beobachteten höheren „Baseline“-ΔIAmi-Werte durch eine
erhöhte Po vor Chymotrypsin-Aktivierung verursacht werden.
Zusammenfassend lässt sich feststellen, dass die Substitution von βV348 durch
andere Aminosäuren in den meisten Fällen die mittlere Po verändert. Nur im
Fall der stark inhibitorischen Mutanten V348Q und V348del wird der „loss-of-
function“-Effekt durch eine reduzierte Expression der Kanäle in der Membran
verursacht.

42
V34
8G
V34
8K
V34
8M
V34
8L
V34
8W
V34
8E wt
V34
8D
0
1
2
3
4
5
6- Chymotrypsin + Chymotrypsin
I A
mi (n
orm
alis
iert
)
N=3
0.5 1.0 1.5 2.02
3
4
5wt
V348M
V348G
V348L
V348W
V348K
V348D
V348E
IAmi (normalisiert)
Rela
tive
Aktivi
eru
ng
A
B
Abb. 18: Aktivierung der unterschiedlichen Mutanten mit Chymotrypsin (mod. nach [55]).
A, Wildtyp-ENaC (wt) und die unterschiedlichen Mutanten wurden in Oozyten exprimiert und
ΔIAmi mit der Zwei-Elektroden-Spannungsklemme detektiert. ENaC wurde mit Chymotrypsin
aktiviert wie in Abb. 12A gezeigt. Die Daten stellen eine Zusammenfassung aus je 47 Oozyten
für Wildtyp- und βV348M-ENaC und je 21-24 Oozyten für die übrigen Mutanten dar. Um die
Variabilität von Batch zu Batch zu korrigieren, wurden die Daten auf den durchschnittlichen
ΔIAmi von Wildtyp exprimierenden Oozyten vor Chymotrypsin-Aktivierung normalisiert. B, Die
gleichen Daten wie in A. Die relative Aktivierung durch Chymotrypsin wurde gegen den auf den
Wildtyp normaliserten ΔIAmi vor Chymotrypsin-Aktivierung aufgetragen.

43
6. Diskussion
Im Rahmen dieser Arbeit wurde die Auswirkung einer Mutation der β-
Untereinheit von ENaC auf die Funktion des Kanals untersucht. Die Mutation
wurde in einem Patienten mit ausgeprägten Symptomen der zystischen Fibrose
entdeckt [51]. Insgesamt stellte sich ein stimulatorischer Effekt auf ΔIami von
etwa 40% heraus. Die funktionelle Charakterisierung wurde in Xenopus laevis
Oozyten als heterologem Expressionssystem durchgeführt.
6.1 Xenopus laevis Oozyten als heterologes Expressions-
system
Die Oozyten bieten wie jedes andere wissenschaftliche Modell Vor- und
Nachteile, so in Bezug auf die Handhabung der Oozyten und die
Interpretierbarkeit der Ergebnisse. Attraktiv machen das System zum einen der
große Durchmesser der Oozyten von etwa 1,1 mm (Stadium V und VI nach
Dumont), was die Handhabung der Zellen erleichtert [21]. Zum anderen weisen
die Oozyten einen gut ausgestatteten Proteinbiosyntheseapparat mit Proteinen,
Enzymen und Organellen auf, der für die embryonale Entwicklung zur
Verfügung steht, aber auch zur Expression exogener mRNA entsprechend
verwendet werden kann [45]. Ebenso laufen in der Oozyte posttranslationale
Modifikationen wie Glykosylierungen, Acetylierungen und Phosphorylierungen
auch von fremden Proteinen in korrekter Weise ab [16]. Die Oozyten sind dazu
befähigt, Proteine auch gemäß ihrer Funktion an die richtige Lokalisation zu
bringen, so dass integrale Membranproteine, wie Ionenkanäle, die
Oozytenmembran erreichen [16]. Dadurch bleiben die Eigenschaften von
Ionenkanälen nahezu unverändert, obwohl sie in fremden Zellen exprimiert
werden. Auch die Tatsache, dass weibliche Frösche das ganze Jahr über eine
Vielzahl von Oozyten aller Entwicklungsstufen beherbergen und auch die
Haltung der Frösche sich weitgehend unproblematisch gestaltet, gehört zu den
Vorteilen des Oozytenexpressionssystems.
Als Nachteile oder Einschränkungen des Systems sind zu nennen, dass Fälle
von nicht korrekt posttranslational modifizierten Proteinen beschrieben sind,
beispielsweise solcher, die sonst gewebespezifisch prozessiert werden [16].
Auch können durch die Defollikulierung bzw. Behandlung mit Kollagenase

44
Defekte in der Oozytenmembran entstehen, die sich ähnlich wie unspezifische
Ionenkanäle verhalten können. Diese können in gleicher Weise wie endogen
exprimierte Kanäle Störquellen für die Messung spezifischer Ströme darstellen
[16].
Fraglich bleibt auch, ob die Ausstattung der second-messenger-Systeme in der
Oozyte der von menschlichen Zellen entspricht, ob folglich Regulationen in
ähnlicher Weise wie im Menschen ablaufen können. Auch die oftmals sehr
heterogenen Ionenkonzentrationen, die sich in Oozyten finden lassen, könnten
für die Messung eine Störquelle darstellen [16, 74].
Generell stellt die heterologe Expression in Oozyten von Xenopus laevis zur
Untersuchung von Ionenkanälen wie ENaC ein gut etabliertes Verfahren dar.
6.2 Einflüsse der cRNA-Qualität
Unterschiede in der cRNA-Qualität können sich auf die Höhe der
Proteinexpression auswirken. Um Fehler zu vermeiden, die auf Unterschieden
der cRNA-Qualität beruhen, wurden in den Versuchen mehrere Batches cRNA
verwendet, um den stimulatorischen Effekt der βV348M-Mutante zu bestätigen.
Auch für alle anderen Mutanten wurden mehrere Batches cRNA angefertigt.
Desweiteren wurde die cRNA der Mutanten jeweils parallel zur entsprechenden
Wildtyp-cRNA synthetisiert, um die Variabilität der Qualität der cRNA möglichst
gering zu halten. Zusätzlich wurden der intrazelluläre und der aus der Membran
stammende Gehalt an β-ENaC von Wildtyp und βV348M-Mutante durch
Western Blot bestimmt, wobei sich eine ähnliche Proteinexpression zeigte.
Aufgrund dieser Maßnahmen ist davon auszugehen, dass der „gain-of-
function“-Effekt der βV348M-Mutation verlässlich nachgewiesen wurde.
Interessanterweise gab es zwei Mutationen (V348Q und V348del), die die
Proteinexpression sowohl intrazellulär als auch in der Zellmembran stark
beeinträchtigten. Als Grund für dieses Ergebnis kommt neben einer im
Vergleich zum Menschen unterschiedlichen Verwendung von bestimmten
Codons in Xenopus laevis Oozyten auch eine mögliche Erkennung als falsch
gefaltetes oder unreifes Protein mit anschließender Degradation in Frage. Der
Nachweis der reduzierten Proteinexpression als Ursache für den „loss-of-
function“-Effekt dieser Mutanten durch Western Blot bestätigt die Validität der in
dieser Arbeit verwendeten Methoden.

45
6.3 Einfluss der intrazellulären Na+-Konzentration
Bei der von Kellenberger et al. untersuchten Liddle-Mutation der β-Untereinheit
konnten in entsprechend injizierten Oozyten nach Inkubation mit Na+-armer
Lösung keine Steigerungen von ΔIAmi im Vergleich zum Wildtyp gefunden
werden [36]. Nach Inkubation in Na+-reicher Lösung und dadurch gesteigerter
intrazellulärer Na+-Konzentration zeigte sich dagegen ein im Vergleich zum
Wildtyp etwa 3-fach erhöhter ΔIAmi der Mutante. Diese Resultate sind Folge der
gestörten „Na+-Feedback-Inhibition“ bei Liddle-Mutanten. Normalerweise führt
die Inkubation in Na+-reicher Lösung (z.B. ND96) in mit ENaC-cRNA injizierten
Oozyten zu einer Reduktion der von ENaC-vermittelten Ströme im Vergleich zu
in Na+-armer Lösung (z.B. ND9) inkubierten Oozyten. Dieses Phänomen wird
durch eine reduzierte Expression der Kanäle in der Oberflächenmembran
verursacht [36, 43, 72]. Auch in den Versuchen dieser Arbeit wurde dieser
Effekt beobachtet. ΔIAmi war bei Wildtyp und βV348M-Mutante nach Inkubation
mit ND96 im Vergleich zur Inkubation mit ND9 stark verringert. Die Reduktion
der Expression von ENaC nach Inkubation in Na+-reicher Lösung fiel bei der
von Kellenberger et al. untersuchten Liddle-Mutante jedoch deutlich geringer
aus [36], was sich in einer Vergrößerung des „gain-of-function“-Effekts unter
diesen Bedingungen zeigte. Eine derartige Steigerung des stimulatorischen
Effekts auf ΔIAmi konnte bei der βV348M-Mutante gegenüber dem Wildtyp-Kanal
nach Inkubation der Oozyten in ND96 nicht gezeigt werden. Diese Ergebnisse
deuten darauf hin, dass bei der βV348M-Mutation im Gegensatz zu der von
Kellenberger et al. untersuchten Liddle-Mutation der Mechanismus der Na+-
Feedback-Inhibition für die Ausbildung des „gain-of-function“-Effekts keine Rolle
spielt. In der Tat bewirkt die Liddle-Mutation überwiegend eine Zunahme der
Oberflächenexpression des Kanals, wohingegen die Befunde dieser Arbeit
zeigen, dass die βV348M-Mutation vor allem die mittlere Po erhöht.
6.4 Mittlere Po
ENaC kann in mindestens zwei Zuständen in der Plasmamembran vorliegen,
nämlich als Kanal mit hoher Offenwahrscheinlichkeit (Po ~0,5), aber auch als
Kanal mit sehr niedriger Po von kleiner 0,05 als sog. „near-silent“-Kanal [9, 38,
58]. Aufgrund dieser Heterogenität der Po von ENaC lässt sich die mittlere Po

46
mit der Patch-Clamp-Technik nur schlecht abschätzen [28]. Zudem können
„near-silent“-Kanäle in der Patch-Clamp-Technik nur schwer erfasst werden
[19]. Die Größe des „gain-of-function“-Effekts der βV348M-Mutante im Vergleich
zum Wildtyp ließ außerdem nur einen geringen Unterschied der mittleren Po
erwarten, so dass für eine verlässliche Detektion dieses Unterschieds mit der
Patch-Clamp-Technik sehr viele Einzelmessungen notwendig gewesen wären.
Deshalb wurde die Bestimmung der Po mithilfe einer indirekten Methode
durchgeführt, der als Mechanismus die maximale Aktivierung von ENaC durch
MTSET zugrunde lag. Hierfür wurden βS520C-Mutanten von Wildtyp und
βV348M-ENaC hergestellt und mit MTSET aktiviert. Diese Methode wurde
bereits für die Untersuchung anderer ENaC-Mutanten erfolgreich eingesetzt
[30, 32].
Die Aktivierung mit MTSET zeigte eine gesteigerte mittlere Po der βV348M-
Mutante, die ausreicht, um den stimulatorischen Effekt der βV348M-Mutante
gegenüber dem Wildtyp vollständig zu erklären.
Falls die Stimulation allein auf einer Erhöhung der Po beruht, sollte die Anzahl
der Kanäle in der Zellmembran unverändert sein. Dies würde sich in ähnlichen
Werten von ΔIAmi des Wildtyps und der βV348M-Mutante nach maximaler
Stimulation mit MTSET widerspiegeln, die in den Experimenten auch tatsächlich
nachgewiesen werden konnten. Durch die unabhängige Methode der
Biotinylierung der Membranproteine konnten zudem gleiche Mengen an βENaC
in der Membran für Wildtyp und βV348M-Mutante nachgewiesen werden. Der
„gain-of-function“-Effekt der βV348M-Mutante beruht deshalb vermutlich allein
auf einer gesteigerten Po bei unveränderter Expression in der Zellmembran.
Ursächlich für die Erhöhung der mittleren Po der βV348M-Mutante könnte zum
einen eine im Vergleich zum Wildtyp geringere Anzahl von „near-silent“-
Kanälen in der Zellmembran sein, während aktive Kanäle entsprechend
vermehrt exprimiert sein könnten. Diese Möglichkeit wurde zum Beispiel bei der
αW493R-Mutante beschrieben [54]. Der stimulatorische Effekt dieser Mutante
belief sich auf einer 4-fachen Steigerung von ΔIAmi. Der im Vergleich dazu nur
geringe Unterschied der mittleren Po zwischen Wildtyp-ENaC und βV348M-
Mutante macht es jedoch unwahrscheinlich, dass die βV348M-Mutation die
mittlere Po durch eine deutliche Reduktion des Anteils an „near-silent“-Kanälen
in der Zellmembran steigert. Daher ist es wahrscheinlicher, dass eine

47
zusätzliche Po-Erhöhung schon aktiver Kanäle für den stimulatorischen Effekt
der βV348M-Mutante verantwortlich ist.
6.5 Proteolytische Kanalaktivierung mit Chymotrypsin
Sowohl endogene als auch exogene Proteasen können ENaC aktivieren, indem
sie den Kanal in der extrazellulären Schleife schneiden. Die Folge ist eine
Erhöhung der mittleren Po und weniger eine Erhöhung der Zahl n der in die
Membran integrierten Kanäle [27].
Serinproteasen, zu denen auch Trypsin und Chymotrypsin gehören, spalten
ENaC während der Kanal-Biosynthese oder an der Zelloberfläche in
extrazellulären Domänen der α- und γ-Untereinheit [33, 60]. Die geschnittenen
Kanäle stellen hierbei vermutlich die reiferen, aktiveren Kanäle dar.
In Oozyten aktiviert Trypsin makroskopische ENaC-Ströme in ähnlicher Weise
wie Chymotrypsin [19, 22, 32, 54]. Zu Beginn der Trypsin-Exposition entstehen
jedoch vorübergehend Calcium-aktivierte Chlorid-Leitfähigkeiten, die störende
einwärts gerichtete Stromkomponenten verursachen. Unter Chymotrypsin
lassen sich ähnliche Effekte nicht erkennen. Deshalb wurde es in der
vorliegenden Arbeit verwendet, um einen Unterschied in der proteolytischen
Aktivierbarkeit zwischen Wildtyp und Mutanten zu überprüfen [15, 19, 22].
Die Versuche der proteolytischen Aktivierung mit Chymotrypsin zeigten
ähnliche Effekte auf ΔIAmi wie die Bestimmung der mittleren Po mit MTSET. Dies
lässt darauf schließen, dass Chymotrypsin wie MTSET die Kanäle vollständig
aktiviert. Im Widerspruch zu diesen Ergebnissen konnte in einer Arbeit von
Diakov et al. [19] ENaC der Spezies Ratte (rENaC) in αβS518CγrENaC
exprimierenden Oozyten (entspricht αβS520CγENaC in humanem ENaC) nach
vorheriger Stimulation mit MTSET durch Trypsin noch weiter aktiviert werden.
Dies könnte daran liegen, dass Trypsin neben aktiven Kanälen auch „near-
silent“-Kanäle aktivieren kann, wie es für Chymotrypsin in „Outside-Out-
Patches“ von ENaC exprimierenden Oozyten gezeigt werden konnte [54].
Bisher war jedoch nicht hinreichend geklärt, ob MTSET ebenso „near-silent“-
Kanäle aktivieren kann. Die Ergebnisse von Diakov et al. [19] lassen vermuten,
dass MTSET ausschließlich aktive Kanäle aktivieren kann, „near-silent“-Kanäle
von der Aktivierung dagegen nicht erfasst werden. Dadurch könnte die mittlere

48
Po aller an der Zelloberfläche befindlichen Kanäle durch die Ermittlung mit
MTSET überschätzt werden.
Die ähnlichen stimulatorischen Effekte von MTSET und Chymotrypsin in der
vorliegenden Arbeit deuten jedoch darauf hin, dass auch MTSET „near-silent“-
Kanäle aktivieren kann. Die Diskrepanz zu den Ergebnissen von Diakov et al.
kann vermutlich durch Spezies-spezifische Unterschiede im Schaltverhalten
(„Gating“) von ENaC erklärt werden [37]. MTSET bindet mutmaßlich nur an die
offene Konformation des Kanals [37, 66]. Falls die Zeiten in offener
Konformation von humanem „near-silent“-ENaC länger sind als von „near-
silent“-ENaC der Ratte, könnte MTSET humanen ENaC besser aktivieren als
ENaC der Ratte. Vergleichende Untersuchungen der Öffnungszeiten von „near-
silent“-Kanälen von humanem ENaC und ENaC der Ratte sind bisher nicht
bekannt und technisch auch nur schwer durchführbar.
6.6 ENaC-Aktivator S3969
Lu et al. [46] fanden heraus, dass sich eine Spleiß-Variante von humanem β-
ENaC, in der die Aminosäure Valin an Position 348 der β-Untereinheit deletiert
war, nicht durch den neuen niedermolekularen ENaC-Aktivator S3969
aktivieren ließ. Somit könnte eine Mutation an dieser Stelle die Aktivierbarkeit
durch S3969 beeinflussen. Aus diesen Gründen wurde Wildtyp- und βV348M-
ENaC im Vergleich mit S3969 aktiviert.
Der stimulatorische Effekt von S3969 auf ΔIami Wildtyp-ENaC exprimierender
Oozyten war geringer als der von Chymotrypsin oder auch als der von MTSET
auf ΔIami von αβS520CγENaC exprimierenden Oozyten.
Außerdem ließ sich ΔIami nach Stimulation mit S3969 durch Chymotrypsin noch
weiter stimulieren. Für die zusätzliche Stimulation konnte zwar keine Signifikanz
nachgewiesen werden, was vermutlich an der geringen Anzahl von 5 Oozyten
pro untersuchter Gruppe lag, ein Trend zeigte sich aber in allen untersuchten
Oozyten. Dies spricht dafür, dass S3969 ENaC nur unvollständig stimulierte.
Der Grund hierfür könnte darin liegen, dass der maximale Effekt des Aktivators
erst bei einer Konzentration von 30 µM erreicht wird [46]. Im Rahmen dieser
Arbeit wurde jedoch eine 10 µM-Lösung verwendet.
Nach Lu et al. [46] ist die Anwesenheit von βV348 essentiell für den
stimulatorischen Effekt von S3969 auf ΔIami. In der vorliegenden Arbeit ließ sich

49
allerdings auch die βV348M-Mutante durch S3969 stimulieren. Die etwas
geringere relative Aktivierung im Vergleich zum Wildtyp lässt sich durch die
höhere mittlere Po der βV348M-Mutante plausibel erklären. Interessanterweise
ließen sich auch drei andere Mutanten, βV348K, βV348W und βV348E durch
S3969 stimulieren (Daten nicht gezeigt). Somit scheint das Valin an Position
348 der β-Untereinheit von ENaC nicht entscheidend für die aktivierende
Funktion von S3969 zu sein, sondern vielmehr die Anwesenheit einer
Aminosäure an dieser Stelle. Die Deletion von βV348 könnte dagegen die
Tertiärstruktur von ENaC derart verändern, dass eine Stimulation durch S3969
unmöglich wird.
6.7 Weitere βV348-Mutanten und ENaC-Modell
Um den molekularen Mechanismus der Po-Erhöhung der βV348M-Mutante zu
untersuchen, wurden Experimente mit verschiedenen Mutanten an Position 348
der β-Untereinheit durchgeführt. Interessanterweise führte der Austausch von
Valin durch einige andere Aminosäuren an dieser Stelle ebenfalls zu einer
Veränderung der ENaC-Ströme. Als Ursache hierfür stellte sich wie bei der
βV348M-Mutante eine Veränderung der mittleren Po bei gleicher Expression
von ENaC in der Zellmembran heraus. Ausnahmen waren die V348Q- und
V348del-Mutante, die keine entsprechende Expression in der Membran zeigten.
Zur Abschätzung der mittleren Po der Mutanten wurde die proteolytische
Aktvierung mit Chymotrypsin herangezogen und analog zur Aktivierung der
βV348M-Mutante mit Chymotrypsin durchgeführt. Der Kanal ließ sich durch
Chymotrypsin in gleicher Weise wie durch MTSET maximal stimulieren und
kann somit zur Bestimmung der Po herangezogen werden. Auf die Bestimmung
der Po durch Stimulation mit MTSET und der damit verbundenen Herstellung
zahlreicher weiterer Mutanten mit zusätzlicher βS520C-Mutation wurde deshalb
verzichtet.
Die Veränderung der Po durch die verschiedenen Mutanten konnte nicht einfach
durch Größe oder Ladung der Seitenkette der stimulierenden Aminosäuren
erklärt werden.
Aus diesem Grund wurde von Prof. Heinrich Sticht an einem zur Kristallstruktur
von ASIC homologen ENaC-Modell [68] die Mikrodomäne um die Aminosäure
V348 der β-Untereinheit im vermutlich geschlossenen Zustand des Kanals

50
analysiert [55]. Die Analyse zeigt, dass V348 nicht nur mit Aminosäuren aus der
eigenen „Palm“-Region, sondern auch mit dem am Verbindungsstück zwischen
„Palm“ und der β-„ball“-Domäne liegenden M90 (Methionin) interagiert (Abb. 2
bzw. 19A). Im Fall der kleinsten Aminosäure Glycin (V348G) lässt das Fehlen
der Seitenkette eine Lücke entstehen. Dadurch kommt es zu einem
Kontaktverlust zu M90, was wiederum die Stabilität der entsprechenden Region
herabsetzt (Abb. 19B). Eine Mutation zu Lysin (V348K) führt dagegen zu einer
räumlichen Kollision mit M90, außerdem zu einer elektrostatischen Abstoßung
von K350 (Lysin) (Abb. 19C). Kollisionen der Aminosäure 348 mit M90 sind in
ähnlicher Weise bei V348M (Abb. 19D), V348L (Leucin) und V348W
(Tryptophan) zu beobachten (Daten nicht gezeigt).
Alle bisher genannten Mutanten sind entweder durch den Bindungsverlust zu
M90 oder durch räumliche Kollisionen mit M90 charakterisiert. Beide Effekte
sind energetisch gesehen unvorteilhaft und destabilisieren vermutlich die
Interaktion von „Palm“-und β-„Ball“-Domäne. Sie erleichtern dadurch vermutlich
den Übergang in die offene Konformation des Kanals. Die auf diese Art
entstandene Erhöhung der Po könnte die Ursache für die ΔIAmi-steigernden
Effekte dieser Mutanten sein. Eine genauere strukturelle Interpretation dieses
Effekts kann jedoch erst nach Vorliegen der Kristallstruktur des offenen Kanals
erfolgen. Diese liegt momentan jedoch noch nicht vor.
Im Gegensatz zu den bisher beschriebenen Mutationen führt die Mutation zu
Glutamat (βV348E) oder Aspartat (βV348D) im Modell zur Ausbildung einer
neuen, stabilisierenden Salzbrücke mit K350 (Abb. 19E). Dieselbe Art von
Interaktion ist interessanterweise auch in der Kristallstruktur des homologen
ASIC 1-Kanals zu beobachten (Abb. 19F). Diese Mutanten stabilisieren durch
die Salzbrücke den geschlossenen Zustand von ENaC. Dies könnte die
Ursache dafür sein, dass die βV348E-Mutante keinen stimulatorischen Effekt
auf ΔIAmi zeigt, die βV348D-Mutante sogar einen leicht inhibitorischen Effekt.
Zusammenfassend stehen die Vorhersagen, die anhand des ENaC-Modells
getroffen wurden, im Einklang mit den experimentellen Befunden der
vorliegenden Doktorarbeit.

51
Abb. 19: Räumliche Effekte verschiedener βV348-Mutationen im ENaC-Modell (mod. nach
[55]). A, Position 348 der β-Untereinheit in der Struktur des Wildtyps: V348 ist grün dargestellt,
die räumlich anliegenden Aminosäuren samt Seitenketten sind im Kalottenmodell abgebildet.
Die Aminosäuren M90 und K350, die für die Interaktionen der Mutanten von Bedeutung sind,
sind markiert. B, Gleiche Ansicht wie in A, jetzt für die V348G-Mutation. Die Lücke, die durch
das Fehlen der Seitenkette von Glycin zwischen G348 (grün) und M90 entsteht, ist gut sichtbar.
C, Die V348K-Mutante resultiert in räumlichen Kollisionen mit M90 (roter Pfeil) und
elektrostatischer Abstoßung von K350 (brauner Pfeil). D, M348 verursacht Kollisionen mit M90
(roter Pfeil). E, D348 bildet eine neue Salzbrücke mit K350 (grüner Pfeil) aus. F, E278-R280
(Glutamat-Arginin) Salzbrücke in der Kristallstruktur des homologen ASIC 1-Ionenkanals.
Weitere benachbarte Aminosäuren von β348 wurden in den Feldern C bis F aus Gründen der
Übersichtlichkeit nicht abgebildet.
A B
D
C
E F

52
6.8 Klinische Relevanz der βV348M-Mutation
Der „gain-of function“-Effekt der βV348M-Mutante konnte auf eine Erhöhung der
Po zurückgeführt werden. Dieser Befund steht in Einklang mit dem
pathophysiologischen Konzept, dass bei betroffenen Patienten durch eine
ENaC-Überfunktion vermehrt Na+-Ionen und dadurch Flüssigkeit im
Respirationstrakt resorbiert werden. Durch Reduktion der ASL-Höhe und der
„mukoziliären Clearance“ könnten dann die typischen pulmonalen Symptome
der CF entstehen. In der Vergangenheit wurden bereits „gain-of-function“-
Mutationen in ENaC beschrieben, die bei Patienten mit CF-ähnlicher
Lungenaffektion entdeckt wurden und dort wahrscheinlich auf ähnliche Weise
pathophysiologisch relevant sind [3, 54, 64].
Die von Azad et al. [3] und Sheridan et al. [64] beschriebenen CF-Patienten
zeigten alle einen milderen Phänotyp. Mutesa et al. berichteten bei dem
Patienten, der die βV348M-Mutation in seinem Genom trägt, jedoch von einer
schweren Ausprägung seiner CF-ähnlichen Symptome [51]. Es ist deshalb
wahrscheinlich, dass neben dem stimulatorischen Effekt der βV348M-Mutation
noch andere Faktoren zum Phänotyp des Patienten beitragen.
Bei dem Patienten wurde zusätzlich eine F693L-CFTR-Mutation gefunden. Die
schwere Ausprägung der Symptome könnte folglich durch einen
synergistischen Effekt zwischen einer ENaC-„gain-of-function“-Mutation und
einer CFTR-„loss-of-function“-Mutation zustande kommen. Die F693L-Mutation
wurde jedoch als ein Polymorphismus beschrieben, von dem im Vergleich zu
Wildtyp-CFTR in COS1-Zellen kein Effekt auf den Reifungsprozess des Kanals
beschrieben wurde. Außerdem konnte im Oozytenexpressionssystem keine
Auswirkung auf die Chloridkanaleigenschaften der F693L-Mutante gezeigt
werden [71]. Aus diesem Grund ist es unwahrscheinlich, dass die F693L-
Mutation direkt krankheitsverursachend ist.
Interessanterweise berichten Mutesa et al. [51] von einem zweiten Patienten mit
schwerwiegenden CF-ähnlichen Symptomen, der neben dieser F693L-CFTR-
Mutation eine βG442V-ENaC-Mutation in seinem Genom trägt. Von der
βG442V-Mutation wurde berichtet, dass sie keine signifikanten Effekte auf
ENaC-vermittelte Na+-Ströme in Oozyten ausübt [2, 53].

53
Dieser Fall zeigt, dass Patienten mit einem CF-Phänotyp Mutationen in ENaC
und CFTR aufweisen können, die in Expressionsmodellen keine Auswirkungen
auf die gemessenen Ströme haben.
Im respiratorischen Epithel interagieren ENaC und CFTR auf komplexe Art und
Weise. Dabei führt die Aktivierung von CFTR zu einer Inhibition von ENaC über
bisher unbekannte Mechanismen [6, 12]. So könnte die F693L-CFTR-Mutation
zu einem Verlust der inhibitorischen Wirkung von CFTR auf ENaC führen und
dadurch zusätzlich die ASL-Höhe durch gesteigerte Na+-Resorption
vermindern.
Diese Überlegungen zeigen, wie komplex und unverstanden die Entstehung
des Lungenphänotyps bei CF ist. Dies wird auch dadurch verdeutlicht, dass
nicht in allen Patienten mit atypischer CF und keinem oder nur einem
betroffenen CFTR-Allel ENaC-Mutationen mit stimulierendem Effekt auf ΔIami im
Oozytenexpressionsmodell nachgewiesen wurden. Andere bei solchen
Patienten identifizierte Mutationen hatten keinen oder sogar inhibitorische
Effekte auf die ENaC-Funktion [3, 32, 64]. Deshalb ist eine im
Expressionssystem nachgewiesene Überfunktion von ENaC sicher nicht in allen
Patienten mit CF-artigen Symptomen ohne typische Mutationen in beiden CFTR
Allelen eine Erklärung für die Krankheitssymptome.
Es ergibt sich die Frage, wie die Regulation der Höhe der ASL in Patienten
ohne CFTR-Mutationen und mit ENaC-Mutationen ohne erkennbaren Effekt auf
die ENaC-Funktion gestört sein kann. Es ist denkbar, dass eine ENaC-Mutation
keinen Effekt auf die Kanalfunktion an sich ausübt, aber den inhibitorischen
Effekt von CFTR auf ENaC vermindert. Dies würde wiederum zu einer
gesteigerten Na+-Resorption führen, die sich aber in einfachen
Expressionsstudien nicht erkennen lässt. Dafür wären aufwändige
Koexpressionsstudien von CFTR mit Wildtyp-ENaC beziehungsweise dem
mutierten ENaC erforderlich. Zusätzlich könnten die ENaC-Mutationen einen
Effekt auf die Produktion, die Reifung oder den Transport von CFTR zur
Zellmembran ausüben, was zu einer verminderten Cl--Sekretion führen würde.
Beides würde letzten Endes in einer Reduktion der ASL resultieren.
Für die Gruppe der inhibitorischen Mutationen von ENaC ist es schwierig, ein
pathophysiologisches Modell abzuleiten. In diesem Zusammenhang sind
Mutationen in Patienten mit Pseudohypoaldosteronismus I (PHA I) zu nennen.
Auch bei ihnen wurde ein CF-ähnlicher Lungenphänotyp beschrieben [31, 48,

54
62]. Im Gegensatz zu CF-Patienten findet man bei manchen PHA I-Patienten
eine gesteigerte ASL-Höhe und eine erhöhte „mukoziliäre Clearance“ [40]. Die
Pathophysiologie dieser Konstellation ist allerdings noch unklar.
Generell könnte die Entwicklung CF-ähnlicher Symptome in der Lunge mit
Störungen der Regulation der ASL-Höhe jeglicher Art zusammenhängen
unabhängig von dem zugrunde liegenden molekularen Mechanismus und den
jeweils daran beteiligten Ionenkanälen. Um dies herauszufinden, ist weiterhin
intensive Forschung auf dem Gebiet der Pathophysiologie der CF notwendig.
7. Schlussfolgerung
Zusammenfassend konnte im Rahmen dieser Arbeit gezeigt werden, dass die
βV348M-Mutation, die in einem Patienten mit schweren CF-ähnlichen
Symptomen gefunden wurde [51], die Aktivität von ENaC über eine erhöhte
mittlere Kanaloffenwahrscheinlichkeit (Po) steigert. Dieses Ergebnis steht mit
der Hypothese im Einklang, dass ein erhöhter Na+-Transport in respiratorischen
Epithelien durch eine vermehrte Flüssigkeitsresorption die „mukoziliäre
Clearance“ reduziert und dadurch CF-ähnliche Symptome in der Lunge
begünstigt. Zusätzlich zu diesem erhöhten Na+-Transport könnten andere
Faktoren zum ausgeprägten Phänotyp des von der Mutation betroffenen
Patienten beitragen. Aus diesem Grund sind weitere Arbeiten notwendig, um
die Rolle von ENaC in der Pathophysiologie der CF zu klären.

55
8. Literaturverzeichnis
[1] Alvarez de la Rosa D., Canessa C. M., Fyfe G. K., Zhang P. (2000) "Structure and regulation of amiloride-sensitive sodium channels". Annu Rev Physiol 62, 573-594
[2] Ambrosius W. T., Bloem L. J., Zhou L., Rebhun J. F., Snyder P. M.,
Wagner M. A., Guo C., Pratt J. H. (1999) "Genetic variants in the epithelial sodium channel in relation to aldosterone and potassium excretion and risk for hypertension". Hypertension 34, 631-637
[3] Azad A. K., Rauh R., Vermeulen F., Jaspers M., Korbmacher J.,
Boissier B., Bassinet L., Fichou Y., des Georges M., Stanke F., De Boeck K., Dupont L., Balascakova M., Hjelte L., Lebecque P., Radojkovic D., Castellani C., Schwartz M., Stuhrmann M., Schwarz M., Skalicka V., de Monestrol I., Girodon E., Ferec C., Claustres M., Tummler B., Cassiman J. J., Korbmacher C., Cuppens H. (2009) "Mutations in the amiloride-sensitive epithelial sodium channel in patients with cystic fibrosis-like disease". Hum Mutat 30, 1093-1103
[4] Barnard E. A., Miledi R., Sumikawa K. (1982) "Translation of
exogenous messenger RNA coding for nicotinic acetylcholine receptors produces functional receptors in Xenopus oocytes". Proc R Soc Lond B Biol Sci 215, 241-246
[5] Boucher R. C. (2004) "New concepts of the pathogenesis of cystic
fibrosis lung disease". Eur Respir J 23, 146-158 [6] Boucher R. C. (2007) "Cystic fibrosis: a disease of vulnerability to airway
surface dehydration". Trends Mol Med 13, 231-240 [7] Burch L. H., Talbot C. R., Knowles M. R., Canessa C. M., Rossier B.
C., Boucher R. C. (1995) "Relative expression of the human epithelial Na+ channel subunits in normal and cystic fibrosis airways". Am J Physiol 269, C511-518
[8] Button B., Boucher R. C. (2008) "Role of mechanical stress in
regulating airway surface hydration and mucus clearance rates". Respir Physiol Neurobiol 163, 189-201
[9] Caldwell R. A., Boucher R. C., Stutts M. J. (2004) "Serine protease
activation of near-silent epithelial Na+ channels". Am J Physiol Cell Physiol 286, C190-194
[10] Canessa C. M. (2007) "Structural biology: unexpected opening". Nature
449, 293-294 [11] Canessa C. M., Schild L., Buell G., Thorens B., Gautschi I.,
Horisberger J. D., Rossier B. C. (1994) "Amiloride-sensitive epithelial

56
Na+ channel is made of three homologous subunits". Nature 367, 463-467
[12] Chambers L. A., Rollins B. M., Tarran R. (2007) "Liquid movement
across the surface epithelium of large airways". Respir Physiol Neurobiol 159, 256-270
[13] Chang S. S., Grunder S., Hanukoglu A., Rosler A., Mathew P. M.,
Hanukoglu I., Schild L., Lu Y., Shimkets R. A., Nelson-Williams C., Rossier B. C., Lifton R. P. (1996) "Mutations in subunits of the epithelial sodium channel cause salt wasting with hyperkalaemic acidosis, pseudohypoaldosteronism type 1". Nat Genet 12, 248-253
[14] Chraibi A., Horisberger J. D. (2002) "Na self inhibition of human
epithelial Na channel: temperature dependence and effect of extracellular proteases". J Gen Physiol 120, 133-145
[15] Chraibi A., Vallet V., Firsov D., Hess S. K., Horisberger J. D. (1998)
"Protease modulation of the activity of the epithelial sodium channel expressed in Xenopus oocytes". J Gen Physiol 111, 127-138
[16] Dascal N. (1987) "The use of Xenopus oocytes for the study of ion
channels". CRC Crit Rev Biochem 22, 317-387 [17] Davies J., Alton E., Griesenbach U. (2005) "Cystic fibrosis modifier
genes". J R Soc Med 98 Suppl 45, 47-54 [18] De Boeck K., Wilschanski M., Castellani C., Taylor C., Cuppens H.,
Dodge J., Sinaasappel M. (2006) "Cystic fibrosis: terminology and diagnostic algorithms". Thorax 61, 627-635
[19] Diakov A., Bera K., Mokrushina M., Krueger B., Korbmacher C.
(2008) "Cleavage in the γ-subunit of the epithelial sodium channel (ENaC) plays an important role in the proteolytic activation of near-silent channels". J Physiol 586, 4587-4608
[20] Duc C., Farman N., Canessa C. M., Bonvalet J. P., Rossier B. C.
(1994) "Cell-specific expression of epithelial sodium channel α, β, and γ subunits in aldosterone-responsive epithelia from the rat: localization by in situ hybridization and immunocytochemistry". J Cell Biol 127, 1907-1921
[21] Dumont J. N. (1972) "Oogenesis in Xenopus laevis (Daudin). I. Stages
of oocyte development in laboratory maintained animals". J Morphol 136, 153-179
[22] Durieux M. E., Salafranca M. N., Lynch K. R. (1994) "Trypsin induces
Ca2+-activated Cl- currents in X. laevis oocytes". FEBS Lett 337, 235-238 [23] Elia G. (2008) "Biotinylation reagents for the study of cell surface
proteins". Proteomics 8, 4012-4024

57
[24] Elkan E. R. (1938) "The Xenopus Pregnancy Test". Br Med J 2, 1253-1274.2
[25] Epple H. J., Amasheh S., Mankertz J., Goltz M., Schulzke J. D.,
Fromm M. (2000) "Early aldosterone effect in distal colon by transcriptional regulation of ENaC subunits". Am J Physiol Gastrointest Liver Physiol 278, G718-724
[26] Farman N., Talbot C. R., Boucher R., Fay M., Canessa C., Rossier B.,
Bonvalet J. P. (1997) "Noncoordinated expression of α-, β-, and γ-subunit mRNAs of epithelial Na+ channel along rat respiratory tract". Am J Physiol 272, C131-141
[27] Gaillard E. A., Kota P., Gentzsch M., Dokholyan N. V., Stutts M. J.,
Tarran R. (2010) "Regulation of the epithelial Na+ channel and airway surface liquid volume by serine proteases". Pflugers Arch 460, 1-17
[28] Garty H., Palmer L. G. (1997) "Epithelial sodium channels: function,
structure, and regulation". Physiol Rev 77, 359-396 [29] Gurdon J. B., Lane C. D., Woodland H. R., Marbaix G. (1971) "Use of
frog eggs and oocytes for the study of messenger RNA and its translation in living cells". Nature 233, 177-182
[30] Haerteis S., Krueger B., Korbmacher C., Rauh R. (2009) "The δ-
subunit of the epithelial sodium channel (ENaC) enhances channel activity and alters proteolytic ENaC activation". J Biol Chem 284, 29024-29040
[31] Hanukoglu A., Bistritzer T., Rakover Y., Mandelberg A. (1994)
"Pseudohypoaldosteronism with increased sweat and saliva electrolyte values and frequent lower respiratory tract infections mimicking cystic fibrosis". J Pediatr 125, 752-755
[32] Huber R., Krueger B., Diakov A., Korbmacher J., Haerteis S.,
Einsiedel J., Gmeiner P., Azad A. K., Cuppens H., Cassiman J. J., Korbmacher C., Rauh R. (2010) "Functional characterization of a partial loss-of-function mutation of the epithelial sodium channel (ENaC) associated with atypical cystic fibrosis". Cell Physiol Biochem 25, 145-158
[33] Hughey R. P., Bruns J. B., Kinlough C. L., Harkleroad K. L., Tong Q.,
Carattino M. D., Johnson J. P., Stockand J. D., Kleyman T. R. (2004) "Epithelial sodium channels are activated by furin-dependent proteolysis". J Biol Chem 279, 18111-18114
[34] Hummler E., Barker P., Gatzy J., Beermann F., Verdumo C., Schmidt
A., Boucher R., Rossier B. C. (1996) "Early death due to defective neonatal lung liquid clearance in αENaC-deficient mice". Nat Genet 12, 325-328

58
[35] Jasti J., Furukawa H., Gonzales E. B., Gouaux E. (2007) "Structure of acid-sensing ion channel 1 at 1.9 Å resolution and low pH". Nature 449, 316-323
[36] Kellenberger S., Gautschi I., Rossier B. C., Schild L. (1998)
"Mutations causing Liddle syndrome reduce sodium-dependent downregulation of the epithelial sodium channel in the Xenopus oocyte expression system". J Clin Invest 101, 2741-2750
[37] Kellenberger S., Gautschi I., Schild L. (2002) "An external site controls
closing of the epithelial Na+ channel ENaC". J Physiol 543, 413-424 [38] Kellenberger S., Schild L. (2002) "Epithelial sodium channel/degenerin
family of ion channels: a variety of functions for a shared structure". Physiol Rev 82, 735-767
[39] Kerem B., Rommens J. M., Buchanan J. A., Markiewicz D., Cox T. K.,
Chakravarti A., Buchwald M., Tsui L. C. (1989) "Identification of the cystic fibrosis gene: genetic analysis". Science 245, 1073-1080
[40] Kerem E., Bistritzer T., Hanukoglu A., Hofmann T., Zhou Z., Bennett
W., MacLaughlin E., Barker P., Nash M., Quittell L., Boucher R., Knowles M. R. (1999) "Pulmonary epithelial sodium-channel dysfunction and excess airway liquid in pseudohypoaldosteronism". N Engl J Med 341, 156-162
[41] Kimura T., Kawabe H., Jiang C., Zhang W., Xiang Y. Y., Lu C., Salter
M. W., Brose N., Lu W. Y., Rotin D. (2011) "Deletion of the ubiquitin ligase Nedd4L in lung epithelia causes cystic fibrosis-like disease". Proc Natl Acad Sci U S A 108 (8), 3216-3221
[42] Knowles M. R., Boucher R. C. (2002) "Mucus clearance as a primary
innate defense mechanism for mammalian airways". J Clin Invest 109, 571-577
[43] Konstas A. A., Shearwin-Whyatt L. M., Fotia A. B., Degger B.,
Riccardi D., Cook D. I., Korbmacher C., Kumar S. (2002) "Regulation of the epithelial sodium channel by N4WBP5A, a novel Nedd4/Nedd4-2-interacting protein". J Biol Chem 277, 29406-29416
[44] Krueger B., Schlotzer-Schrehardt U., Haerteis S., Zenkel M.,
Chankiewitz V. E., Amann K. U., Kruse F. E., Korbmacher C. (2012) "Four subunits (αβγδ) of the epithelial sodium channel (ENaC) are expressed in the human eye in various locations". Invest Ophthalmol Vis Sci 53, 596-604
[45] Lane C. D. (1981) "The fate of foreign proteins introduced in Xenopus
oocytes". Cell 24, 281-282 [46] Lu M., Echeverri F., Kalabat D., Laita B., Dahan D. S., Smith R. D., Xu
H., Staszewski L., Yamamoto J., Ling J., Hwang N., Kimmich R., Li P., Patron E., Keung W., Patron A., Moyer B. D. (2008) "Small

59
molecule activator of the human epithelial sodium channel". J Biol Chem 283, 11981-11994
[47] Mall M., Grubb B. R., Harkema J. R., O'Neal W. K., Boucher R. C.
(2004) "Increased airway epithelial Na+ absorption produces cystic fibrosis-like lung disease in mice". Nat Med 10, 487-493
[48] Marthinsen L., Kornfalt R., Aili M., Andersson D., Westgren U.,
Schaedel C. (1998) "Recurrent Pseudomonas bronchopneumonia and other symptoms as in cystic fibrosis in a child with type I pseudohypoaldosteronism". Acta Paediatr 87, 472-474
[49] Matsui H., Grubb B. R., Tarran R., Randell S. H., Gatzy J. T., Davis C.
W., Boucher R. C. (1998) "Evidence for periciliary liquid layer depletion, not abnormal ion composition, in the pathogenesis of cystic fibrosis airways disease". Cell 95, 1005-1015
[50] Mennitt P. A., Wade J. B., Ecelbarger C. A., Palmer L. G., Frindt G.
(1997) "Localization of ROMK channels in the rat kidney". J Am Soc Nephrol 8, 1823-1830
[51] Mutesa L., Azad A. K., Verhaeghe C., Segers K., Vanbellinghen J. F.,
Ngendahayo L., Rusingiza E. K., Mutwa P. R., Rulisa S., Koulischer L., Cassiman J. J., Cuppens H., Bours V. (2009) "Genetic analysis of Rwandan patients with cystic fibrosis-like symptoms: identification of novel cystic fibrosis transmembrane conductance regulator and epithelial sodium channel gene variants". Chest 135, 1233-1242
[52] Orenstein D. M., Winnie G. B., Altman H. (2002) "Cystic fibrosis: a
2002 update". J Pediatr 140, 156-164 [53] Persu A., Barbry P., Bassilana F., Houot A. M., Mengual R.,
Lazdunski M., Corvol P., Jeunemaitre X. (1998) "Genetic analysis of the β subunit of the epithelial Na+ channel in essential hypertension". Hypertension 32, 129-137
[54] Rauh R., Diakov A., Tzschoppe A., Korbmacher J., Azad A. K.,
Cuppens H., Cassiman J. J., Dotsch J., Sticht H., Korbmacher C. (2010) "A mutation of the epithelial sodium channel associated with atypical cystic fibrosis increases channel open probability and reduces Na+ self inhibition". J Physiol 588, 1211-1225
[55] Rauh R., Soell D., Haerteis S., Diakov A., Nesterov V., Krueger B.,
Sticht H., Korbmacher C. (2013) "A mutation in the β-subunit of ENaC identified in a patient with cystic fibrosis-like symptoms has a gain-of-function effect". Am J Physiol Lung Cell Mol Physiol 304, L43-55
[56] Renard S., Lingueglia E., Voilley N., Lazdunski M., Barbry P. (1994)
"Biochemical analysis of the membrane topology of the amiloride-sensitive Na+ channel". J Biol Chem 269, 12981-12986

60
[57] Riordan J. R., Rommens J. M., Kerem B., Alon N., Rozmahel R., Grzelczak Z., Zielenski J., Lok S., Plavsic N., Chou J. L., et al. (1989) "Identification of the cystic fibrosis gene: cloning and characterization of complementary DNA". Science 245, 1066-1073
[58] Rossier B. C. (2002) "Hormonal regulation of the epithelial sodium
channel ENaC: N or Po?". J Gen Physiol 120, 67-70 [59] Rossier B. C., Pradervand S., Schild L., Hummler E. (2002) "Epithelial
sodium channel and the control of sodium balance: interaction between genetic and environmental factors". Annu Rev Physiol 64, 877-897
[60] Rossier B. C., Stutts M. J. (2009) "Activation of the epithelial sodium
channel (ENaC) by serine proteases". Annu Rev Physiol 71, 361-379 [61] Roudier-Pujol C., Rochat A., Escoubet B., Eugene E., Barrandon Y.,
Bonvalet J. P., Farman N. (1996) "Differential expression of epithelial sodium channel subunit mRNAs in rat skin". J Cell Sci 109 ( Pt 2), 379-385
[62] Schaedel C., Marthinsen L., Kristoffersson A. C., Kornfalt R.,
Nilsson K. O., Orlenius B., Holmberg L. (1999) "Lung symptoms in pseudohypoaldosteronism type 1 are associated with deficiency of the α-subunit of the epithelial sodium channel". J Pediatr 135, 739-745
[63] Sheppard D. N., Welsh M. J. (1999) "Structure and function of the
CFTR chloride channel". Physiol Rev 79, S23-45 [64] Sheridan M. B., Fong P., Groman J. D., Conrad C., Flume P., Diaz R.,
Harris C., Knowles M., Cutting G. R. (2005) "Mutations in the beta-subunit of the epithelial Na+ channel in patients with a cystic fibrosis-like syndrome". Hum Mol Genet 14, 3493-3498
[65] Shimkets R. A., Warnock D. G., Bositis C. M., Nelson-Williams C.,
Hansson J. H., Schambelan M., Gill J. R., Jr., Ulick S., Milora R. V., Findling J. W., et al. (1994) "Liddle's syndrome: heritable human hypertension caused by mutations in the β subunit of the epithelial sodium channel". Cell 79, 407-414
[66] Snyder P. M., Bucher D. B., Olson D. R. (2000) "Gating induces a
conformational change in the outer vestibule of ENaC". J Gen Physiol 116, 781-790
[67] Stanke F., Becker T., Cuppens H., Kumar V., Cassiman J. J., Jansen
S., Radojkovic D., Siebert B., Yarden J., Ussery D. W., Wienker T. F., Tummler B. (2006) "The TNFalpha receptor TNFRSF1A and genes encoding the amiloride-sensitive sodium channel ENaC as modulators in cystic fibrosis". Hum Genet 119, 331-343
[68] Stockand J. D., Staruschenko A., Pochynyuk O., Booth R. E.,
Silverthorn D. U. (2008) "Insight toward epithelial Na+ channel

61
mechanism revealed by the acid-sensing ion channel 1 structure". IUBMB Life 60, 620-628
[69] Sumikawa K., Houghton M., Emtage J. S., Richards B. M., Barnard
E. A. (1981) "Active multi-subunit ACh receptor assembled by translation of heterologous mRNA in Xenopus oocytes". Nature 292, 862-864
[70] Talbot C. L., Bosworth D. G., Briley E. L., Fenstermacher D. A.,
Boucher R. C., Gabriel S. E., Barker P. M. (1999) "Quantitation and localization of ENaC subunit expression in fetal, newborn, and adult mouse lung". Am J Respir Cell Mol Biol 20, 398-406
[71] Vankeerberghen A., Wei L., Jaspers M., Cassiman J. J., Nilius B.,
Cuppens H. (1998) "Characterization of 19 disease-associated missense mutations in the regulatory domain of the cystic fibrosis transmembrane conductance regulator". Hum Mol Genet 7, 1761-1769
[72] Volk T., Konstas A. A., Bassalay P., Ehmke H., Korbmacher C.
(2004) "Extracellular Na+ removal attenuates rundown of the epithelial Na+-channel (ENaC) by reducing the rate of channel retrieval". Pflugers Arch 447, 884-894
[73] Waldmann R., Champigny G., Bassilana F., Voilley N., Lazdunski M.
(1995) "Molecular cloning and functional expression of a novel amiloride-sensitive Na+ channel". J Biol Chem 270, 27411-27414
[74] Weber W. M. (1999) "Endogenous ion channels in oocytes of xenopus
laevis: recent developments". J Membr Biol 170, 1-12 [75] Xu J. Z., Hall A. E., Peterson L. N., Bienkowski M. J., Eessalu T. E.,
Hebert S. C. (1997) "Localization of the ROMK protein on apical membranes of rat kidney nephron segments". Am J Physiol 273, F739-748
[76] Zhou Z., Treis D., Schubert S. C., Harm M., Schatterny J., Hirtz S.,
Duerr J., Boucher R. C., Mall M. A. (2008) "Preventive but not late amiloride therapy reduces morbidity and mortality of lung disease in βENaC-overexpressing mice". Am J Respir Crit Care Med 178, 1245-1256

62
9. Abbildungsverzeichnis
Abb. 1: Struktur von ENaC in der Zellmembran. 6
Abb. 2: ENaC-Modell. 7
Abb. 3: Von ENaC vermittelter Na+-Transport im ASDN. 8
Abb. 4: Regulation und Fehlregulation des ASL. 10
Abb. 5: Aminosäuresequenzen verschiedener Spezies der Region um βV348.
12
Abb. 6: Xenopus laevis, der afrikanische Krallenfrosch. 13
Abb. 7: Xenopus laevis Oozyten im Reifestadium V und VI nach Dumont. 14
Abb. 8: Schematischer Aufbau der Zwei-Elektroden-Spannungsklemme. 19
Abb. 9: Die βV348M-Mutation stimuliert den ENaC-vermittelten Na+- Strom.
25
Abb. 10: Der stimulatorische Effekt der βV348M-Mutation ist vom Na+- Gehalt der Oozyten unabhängig.
26
Abb. 11: Die βV348M-Mutation erhöht die mittlere Po von ENaC. 29
Abb. 12: Chymotrypsin stimuliert αβγENaC mehr als αβV348MγENaC. 31
Abb. 13: Die βV348M-Mutante wird durch S3969 weniger aktiviert als der Wildtyp.
33
Abb. 14: Chymotrypsin aktiviert Wildtyp-ENaC stärker als S3969. 35
Abb. 15: Aminosäuren, die durch positionsgerichtete Mutagenese zur Generation verschiedener Mutanten eingesetzt wurden.
36
Abb. 16: Aminosäure-Mutationen zeigen unterschiedliche Effekte auf die ENaC-Funktion.
37
Abb. 17: Auswirkungen der unterschiedlichen Mutationen auf die βENaC-Expression intrazellulär und in der Zellmembran.
39
Abb. 18: Aktivierung der unterschiedlichen Mutanten mit Chymotrypsin. 42
Abb. 19: Räumliche Effekte verschiedener βV348-Mutationen im ENaC-Modell.
51
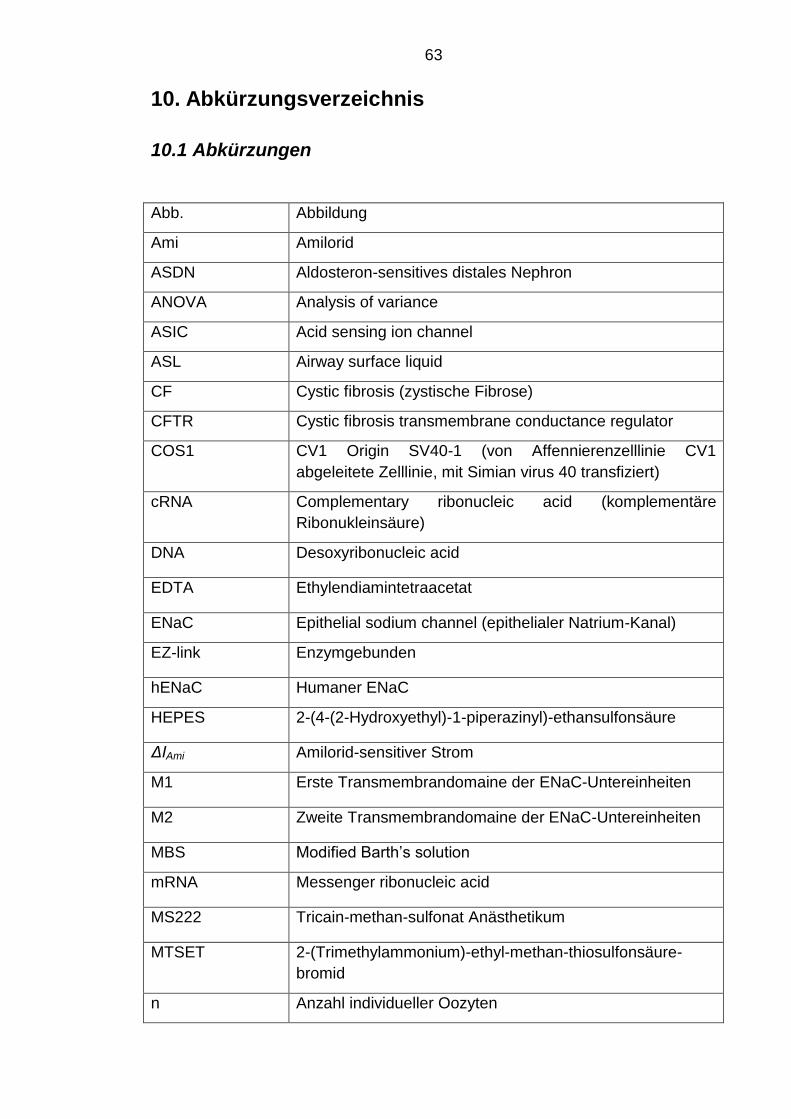
63
10. Abkürzungsverzeichnis
10.1 Abkürzungen
Abb. Abbildung
Ami Amilorid
ASDN Aldosteron-sensitives distales Nephron
ANOVA Analysis of variance
ASIC Acid sensing ion channel
ASL Airway surface liquid
CF Cystic fibrosis (zystische Fibrose)
CFTR Cystic fibrosis transmembrane conductance regulator
COS1 CV1 Origin SV40-1 (von Affennierenzelllinie CV1
abgeleitete Zelllinie, mit Simian virus 40 transfiziert)
cRNA Complementary ribonucleic acid (komplementäre
Ribonukleinsäure)
DNA Desoxyribonucleic acid
EDTA Ethylendiamintetraacetat
ENaC Epithelial sodium channel (epithelialer Natrium-Kanal)
EZ-link Enzymgebunden
hENaC Humaner ENaC
HEPES 2-(4-(2-Hydroxyethyl)-1-piperazinyl)-ethansulfonsäure
ΔIAmi Amilorid-sensitiver Strom
M1 Erste Transmembrandomaine der ENaC-Untereinheiten
M2 Zweite Transmembrandomaine der ENaC-Untereinheiten
MBS Modified Barth’s solution
mRNA Messenger ribonucleic acid
MS222 Tricain-methan-sulfonat Anästhetikum
MTSET 2-(Trimethylammonium)-ethyl-methan-thiosulfonsäure-
bromid
n Anzahl individueller Oozyten

64
N Anzahl Oozyten-Batches
Na+-K+-ATPase Aktive Natrium-Kalium-Pumpe
ND9 Oozytensuperfusionslösung mit 9 mmol/l NaCl
ND96 Oozytensuperfusionslösung mit 96 mmol/l NaCl
Nedd4-2 Neuronal precursor cell-expressed developmentally down-
regulated protein 4-2
NMDG N-methyl-D-glutamat
OR2 Oozyten-Ringer Lösung 2
PHA1 Pseudohypoaldosteronismus Typ 1
PCL Periciliary layer
Po Offenwahrscheinlichkeit
rENaC Epithelial sodium channel (epithelialer Natrium-Kanal) der
Spezies Ratte
RNA Ribonucleic acid
ROMK Renal outer medullary K+-channel
S3969 ENaC-Aktivator
SDS-PAGE Natriumdodecylsulfat-Polyacrylamidgelelektrophorese
SEM Standard error of the mean (Standardfehler)
sulfo-NHS-SS-
Biotin
Sulfosuccinimidyl-2-(biotinamido)-ethyl-1,3'-dithiopropionat
Tris Tris[hydroxymethyl]aminomethan
wt Wildtyp

65
10.2 Nomenklatur der Aminosäuren
Aminosäure Einbuchstabencode
Aspartat D
Glutamat E
Glutamin Q
Glycin G
Leucin L
Lysin K
Methionin M
Tryptophan T
Valin V
10.3 Einheiten
Einheit Bedeutung
A Ampère
°C Grad Celsius
g Gramm
g Mittlere Erdbeschleunigung (G=9,81 m/s2)
h Stunde
l Liter
m Meter
M Molar (mol/l)
mol Mol
Ω Ohm
U Units (Einheiten enzymatischer Aktivität)
V Volt

66
11. Vorveröffentlichungen
Soell D., Korbmacher C., Rauh R. (2010) ”A mutation in the β-subunit of the epithelial sodium channel (ENaC) identified in a patient with atypical cystic fibrosis has a gain-of-function effect”. Acta Physiol, Supp. 677, 198, Abstract P-TUE-24
Rauh R., Soell D., Haerteis S., Diakov A., Nesterov V., Krueger B., Sticht H., Korbmacher C. (2013) "A mutation in the β-subunit of ENaC identified in a patient with cystic fibrosis-like symptoms has a gain-of-function effect". Am J Physiol Lung Cell Mol Physiol 304, L43-55

67
12. Anhang
Primersequenzen der Mutanten:
V348G (vorwärts): 5' CATCGGGGTACTCGGAGACAAGCTTCAGCG 3'
V348G (rückwärts): 5' CGCTGAAGCTTGTCTCCGAGTACCCCGATG 3'
V348K (vorwärts): 5'CGTCCATCGGGGTACTCAAAGACAAGCTTCAGCGCATG 3'
V348K (rückwärts): 5'CATGCGCTGAAGCTTGTCTTTGAGTACCCCGATGGACG 3'
V348M (vorwärts): 5'-CCATCGGGGTACTCATGGACAAGCTTCAGC-3'
V348M (rückwärts): 5'-GCTGAAGCTTGTCCATGAGTACCCCGATGG-3'
V348L (vorwärts): 5' CATCGGGGTACTCCTGGACAAGCTTCAG 3'
V348L (rückwärts): 5' CTGAAGCTTGTCCAGGAGTACCCCGATG 3'
V348W (vorwärts): 5' CCATCGGGGTACTCTGGGACAAGCTTCAGC 3'
V348W (rückwärts): 5' GCTGAAGCTTGTCCCAGAGTACCCCGATGG 3'
V348E (vorwärts): 5' CCATCGGGGTACTCGAAGACAAGCTTCAGCGC 3'
V348E (rückwärts): 5' GCGCTGAAGCTTGTCTTCGAGTACCCCGATGG 3'
V348D (vorwärts): 5' CATCGGGGTACTCGATGACAAGCTTCAGCGC 3'
V348D (rückwärts): 5' GCGCTGAAGCTTGTCATCGAGTACCCCGATG 3'
V348Q (vorwärts): 5' CCATCGGGGTACTCCAGGACAAGCTTCAGC 3'
V348Q (rückwärts): 5' GCTGAAGCTTGTCCTGGAGTACCCCGATGG 3'
V348del (vorwärts): 5'-CATCGGGGTACTCGACAAGCTTCAG-3'
V348del (rückwärts): 5' CTGAAGCTTGTCGAGTACCCCGATG 3'
S520C (vorwärts): 5'CGTCTGGCTGCTCTGCAATCTGGGTGGCCAG3'
S520C (rückwärts):5'CTGGCCACCCAGATTGCAGAGCAGCCAGACG3'

68
13. Danksagung
Ich danke meinem Doktorvater Prof. Dr. med. Christoph Korbmacher, dass er
mir diese Doktorarbeit an seinem Institut ermöglicht hat. Außerdem möchte ich
mich für die hervorragende Betreuung während meiner Zeit in der Physiologie
bedanken, was für mich nicht selbstverständlich ist.
Weiterhin möchte ich meinem Betreuer PD Dr. med. Robert Rauh für seine
große Hilfsbereitschaft und Geduld danken, mit denen er mir mit Rat und Tat
bei der Durchführung und Auswertung der Experimente zur Seite stand. Danke
für die immer zeitnahen Antworten auf aufkommende Fragen, für den
menschlichen Umgang und Gespräche über den Horizont der Arbeit hinaus.
Auch das ist für mich alles andere als selbstverständlich.
Vielen Dank an Sonja Mayer, Celine Grüninger, Jessica Rinke und Dipl.-Biol.
Ralf Rinke für zahlreiche Frosch-Operationen, für das geduldige Anleiten von
neuen Arbeitsmethoden, für etliche cRNA-Herstellungen, für das
selbstverständliche Beantworten aller aufkommenden Fragen, für die
Hilfsbereitschaft, wenn etwas nicht so lief wie gewünscht und für viele lustige
Praktikumsstunden.
Außerdem möchte ich mich für die gute Arbeitsatmosphäre am Institut
bedanken, ohne die mir die Arbeit nur halb so viel Spaß gemacht hätte. Dazu
haben auch Dr. Viatcheslav Nesterov, Dr. Alexej Diakov, Dr. rer. nat. Elena
Rudakova, Prof. Dr. med. Tillmann Volk, Dr. rer. nat. Bettina Krüger, Dr. rer.
nat. Marko Bertog, Dr. med. Michael Wagner, Dr. rer. nat. Silke Härteis und
Dipl.-Biol. Regina Nacken entscheidend dazu beigetragen, nicht zu vergessen
Claudia Ebner, Claudia Großkopf und Axel Günther.
Ein besonderer Dank gilt auch den anderen Doktoranden, die zur gleichen Zeit
Versuche am Institut durchführten. Insbesondere möchte ich mich bei
Christopher Friedrich bedanken, ohne den ich zur Durchführung meiner
Doktorarbeit nicht am physiologischen Institut gelandet wäre und durch den so
mancher langer Labortag kurzweiliger wurde. Danke für die schöne Zeit.

69
Ein großer Dank geht an meine Eltern Renate und Friedrich, an meine Frau
Veronika, meine Geschwister Dorothee, Tabea und Lena und an meine
gesamte Familie mit Freunden, die mich bei der Durchführung dieser Arbeit
unterstützt, ermutigt und in manchem Augenblick auch ertragen haben. Ohne
sie wäre diese Arbeit so nicht möglich gewesen.
Zuletzt möchte ich mich bei Gott bedanken, der mich durch Höhen und Tiefen,
die mit dieser Arbeit verbunden waren, getragen hat.

70
14. Lebenslauf
Persönliche Daten Name Daniel Söll
Geburtsdatum 30. August 1985
Geburtsort Bayreuth
Eltern Renate Söll, geb. Dippon Dr. med. Friedrich Söll
Geschwister Dorothee Langenbucher, geb. Söll Dipl.-Psych. Tabea Söll Lena Söll
Familienstand Verheiratet mit Veronika Söll, geb. Reichstein
Schulische Ausbildung 1991 – 1995 Volksschule St. Johannis in Bayreuth
1995 – 2004 Markgräfin-Wilhelmine-Gymnasium in Bayreuth
06/2004 Allgemeine Hochschulreife Note 1,0
Tätigkeiten zwischen Abitur und Beginn des Medizinstudiums 09/2004 – 04/2005 Englischsprachige Bibelschule der Fackelträger
„Bodenseehof“ in Fischbach/Friedrichshafen
06/2005-07/2005 Krankenpflegepraktikum im Stadtkrankenhaus Pegnitz (Innere Medizin)
Hochschulstudium 10/2005 - 11/2011 Studium der Humanmedizin an der FAU Erlangen-
Nürnberg
09/2007 Erster Teil der Ärztlichen Prüfung. Note: „sehr gut“ (1,5)
10/2011 Zweiter Teil der Ärztlichen Prüfung Note: „sehr gut“ (1,0)
11/2011 Approbation als Arzt

71
Praktika und Famulaturen 03/2006 Krankenpflegepraktikum im Klinikum Kulmbach (Unfallchirurgie)
03/2007 Famulatur in der Sana Klinik Pegnitz Gastroenterologie und Kardiologie
03/2009-04/2009 Famulatur am Klinikum Nürnberg Nord Anästhesiologie
09/2009-10/2009 Famulatur am Klinikum am Europakanal Erlangen Neurologie und neurologische Rehabilitation
Famulaturen im ambulanten Bereich:
02/2010-03/2010 Gemeinschaftspraxis Dr. med. Leppik und Dr. med. Eberhardt, Erlangen
Kinder- und Jugendmedizin
03/2010 Praxis Dr. med. Friedrich Söll, Bayreuth Allgemeinmedizin Praktisches Jahr 08/2010-12/2010 Innere Medizin am Klinikum Bayreuth Fachbereiche: Nephrologie und Kardiologie
12/2010-03/2011 Anästhesiologie am Universitätsklinikum Erlangen
Einsatzkliniken: Neurochirurgie/Ophthalmologie Gynäkologie
Zahn-, Mund- und Kieferchirurgie
03/2011-07/2011 Chirurgie am Klinikum Bayreuth Fachbereiche: Viszeralchirurgie und Unfallchirurgie Promotion Seit 01/09 Doktorvater: Prof. Dr. med. Korbmacher,
Institut für zelluläre und molekulare Physiologie, FAU Erlangen-Nürnberg
Tätigkeit als Tutor am Institut für Zelluläre und Molekulare Physiologie (4 Semester)
05/2009-07/2009 Stipendium des SFB 423 (Collaborative Research Center, Kidney injury: pathogenesis and regenerative mechanisms, FAU Erlangen-Nürnberg)
04/09-08/10 Teilnahme am Doktorandenprogramm des SFB 423

72
Kongressteilnahmen 03/2010 Postervorstellung: Joint Meeting of the
Scandinavian and German Physiological Societies, Kopenhagen, Dänemark
07/2010 Posterpreis: Summer School der
Doktorandenprogramme SFB 423 Universität Erlangen/SFB 699 Universität Regensburg, Hersbruck
10/2010 Postervorstellung: 3rd International SFB 423
Symposium "Molecular Targets in Renal Disease", Bamberg
Thema jeweils:
A mutation in the β-subunit of the epithelial sodium channel (ENaC) identified in a patient with atypical cystic fibrosis has a gain-of-function effect
D. Soell, C. Korbmacher, R. Rauh Aktuelle Beschäftigung Seit 04/12 Assistenzarzt Innere Medizin, Klinikum Bamberg,
Medizinische Klinik III (Nephrologie, Rheumatologie, Osteologie), Chefarzt PD Dr. med. Clemens Grupp
Stand Januar 2013


















