
Atualização das exigências de aminoácidos na nutrição de...
14
29ª Reunião do CBNA – Congresso sobre Nutrição de Aves e Suínos 2015 De 02 a 04 de dezembro de 2015 – Hotel Fonte Colina Verde – São Pedro, SP Atualização das exigências de aminoácidos na nutrição de matrizes Gustavo Gattás 1 , Cesar Augusto Pospissil Garbossa, Maíra Resende, Vinícius de Souza Cantarelli 2 1 AnimalNutri Ciência e Tecnologia 2 Departamento de Zootecnia, Universidade Federal de Lavras - UFLA 1. Introdução O conceito de que os animais necessitam de proteína, substâncias orgânicas indispensáveis para o desenvolvimento adequado e mantença, foi reconhecido há aproximadamente 200 anos atrás (Carpenter, 2003a; Munro, 1964). Desde então, o nosso entendimento das exigências de proteína/aminoácidos e a dinâmica do metabolismo proteico, incluindo a biossíntese e degradação, cresceu consideravelmente (Carpenter, 2003b; Munro, 1964). O melhoramento genético na suinocultura buscou animais de alta produção. Considerando as matrizes suínas, a maior produtividade foi direcionada para o maior número de leitões. No entanto, este incremento é associado a um menor peso ao nascimento e maior variabilidade entre estes. Somado a isto a produção de leite também foi melhorada, no entanto, as fêmeas não têm capacidade de consumo suficiente para suprir suas exigências, levando a maior mobilização dos tecidos corporais (Jones & Stahly, 1999). A mobilização excessiva durante a lactação leva ao aumento do intervalo desmame estro (geralmente não inferior a 10 dias), reduz a taxa de concepção e aumenta a taxa de mortalidade embrionária (Close & Mullan, 1996). Por isso, o programa nutricional de matrizes em gestação deve prezar pelo fornecimento de nutrientes de forma a atender à exigência adequada para formação e desenvolvimento dos conceptos, placenta, anexos embrionários e glândula mamária. Já na fase de lactação, o programa nutricional deve priorizar tanto a exigência para produção de leite bem como adequar-se de modo que a mobilização corporal não seja demasiada e assim, possibilite condição adequada para os partos subsequentes (Hauang et al., 2013). Dessa forma, a potencialização de utilização da porção proteica se faz importante para o sucesso do sistema produtivo, visto que esta tem alto impacto sobre o custo das dietas. Os aminoácidos, além da contribuição para deposição de tecido muscular, exercem funções metabólicas importantes, cabendo ao nutricionista adequar as dietas das matrizes de acordo com o mercado em que está inserido. Assim, o objetivo desta palestra é atualizar as exigências nutricionais de aminoácidos na nutrição de matrizes suínas.
Transcript of Atualização das exigências de aminoácidos na nutrição de...

29ª Reunião do CBNA – Congresso sobre Nutrição de Aves e Suínos 2015 De 02 a 04 de dezembro de 2015 – Hotel Fonte Colina Verde – São Pedro, SP
Atualização das exigências de aminoácidos na nutrição de matrizes
Gustavo Gattás1, Cesar Augusto Pospissil Garbossa, Maíra Resende, Vinícius de Souza
Cantarelli2
1AnimalNutri Ciência e Tecnologia 2Departamento de Zootecnia, Universidade Federal de Lavras - UFLA
1. Introdução
O conceito de que os animais necessitam de proteína, substâncias orgânicas indispensáveis para o
desenvolvimento adequado e mantença, foi reconhecido há aproximadamente 200 anos atrás (Carpenter,
2003a; Munro, 1964). Desde então, o nosso entendimento das exigências de proteína/aminoácidos e a
dinâmica do metabolismo proteico, incluindo a biossíntese e degradação, cresceu consideravelmente
(Carpenter, 2003b; Munro, 1964).
O melhoramento genético na suinocultura buscou animais de alta produção. Considerando as
matrizes suínas, a maior produtividade foi direcionada para o maior número de leitões. No entanto, este
incremento é associado a um menor peso ao nascimento e maior variabilidade entre estes. Somado a isto a
produção de leite também foi melhorada, no entanto, as fêmeas não têm capacidade de consumo suficiente
para suprir suas exigências, levando a maior mobilização dos tecidos corporais (Jones & Stahly, 1999). A
mobilização excessiva durante a lactação leva ao aumento do intervalo desmame estro (geralmente não
inferior a 10 dias), reduz a taxa de concepção e aumenta a taxa de mortalidade embrionária (Close & Mullan,
1996).
Por isso, o programa nutricional de matrizes em gestação deve prezar pelo fornecimento de
nutrientes de forma a atender à exigência adequada para formação e desenvolvimento dos conceptos,
placenta, anexos embrionários e glândula mamária. Já na fase de lactação, o programa nutricional deve
priorizar tanto a exigência para produção de leite bem como adequar-se de modo que a mobilização corporal
não seja demasiada e assim, possibilite condição adequada para os partos subsequentes (Hauang et al.,
2013).
Dessa forma, a potencialização de utilização da porção proteica se faz importante para o sucesso
do sistema produtivo, visto que esta tem alto impacto sobre o custo das dietas. Os aminoácidos, além da
contribuição para deposição de tecido muscular, exercem funções metabólicas importantes, cabendo ao
nutricionista adequar as dietas das matrizes de acordo com o mercado em que está inserido. Assim, o
objetivo desta palestra é atualizar as exigências nutricionais de aminoácidos na nutrição de matrizes suínas.

29ª Reunião do CBNA – Congresso sobre Nutrição de Aves e Suínos 2015 De 02 a 04 de dezembro de 2015 – Hotel Fonte Colina Verde – São Pedro, SP
2. Gestação
2.1 Dinâmica dos aminoácidos durante a gestação
As exigências nutricionais de uma matriz suína em gestação podem ser divididas em diferentes
componentes de acordo com a sua necessidade fisiológica: mantença, crescimento maternal (mais
expressivo em marrãs) e reprodução (estruturas relacionadas [útero, placenta, glândula mamária], e
crescimento fetal). A exigência de cada elemento depende das diferentes fases e dos fenômenos metabólicos
que incidem sobre o período gestacional, bem como sobre a ordem de parto que a fêmea se encontra.
Neste sentido, alimentar as fêmeas em gestação com um nível de aminoácidos fixos durante a
gestação é incorreto, pois se assumiria que a exigência de nutrientes possui uma distribuição igual durante
todo o período gestacional. Dessa forma, o foco metabólico de recuperação da condição da fêmea gestante
pós desmame até a taxa de síntese fetal no final da gestação é alterado. O peso fetal, o conteúdo proteico
fetal e mamário aumentam em 5, 18 e 27 vezes, respectivamente, nos últimos 45 dias de gestação
(McPherson et al., 2004; Ji et al., 2006), assim, esse grande incremento no peso fetal e ganho proteico
indicam que a exigência aminoacídica deve ser maior na fase final de gestação comparada a fase inicial.
Apesar dos níveis de aminoácidos utilizados por pesquisadores e as formas de avaliação divergirem
(modelagem ou experimentação animal), todos concordam que exigência aminoacídica possui incremento
ao final da gestação (Moehn e Ball, 2013). Desta forma, propõe-se o fornecimento de nutrientes em uma
forma em “U”, também conhecida como “high-low-high” durante a gestação. Este manejo pode ser
realizado pelo aumento da quantidade de ração fornecida ou de uma forma mais adequada, pelo
fornecimento de rações com diferentes concentrações nutricionais.
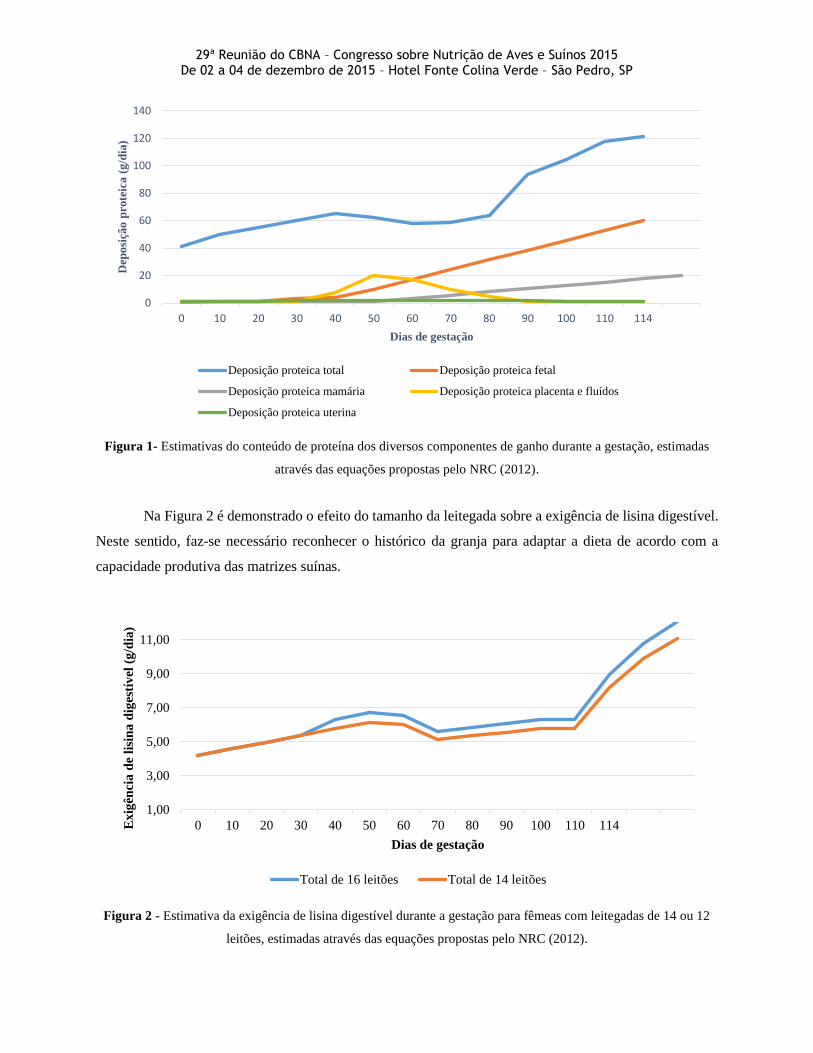
Para ilustrar a importância deste fornecimento diferenciado durante a fase de gestação, na Figura 1
segue o exemplo de deposição proteica nos diferentes componentes das fêmeas durante a gestação através
das equações propostas pelo NRC 2012.
29ª Reunião do CBNA – Congresso sobre Nutrição de Aves e Suínos 2015 De 02 a 04 de dezembro de 2015 – Hotel Fonte Colina Verde – São Pedro, SP
Figura 1- Estimativas do conteúdo de proteína dos diversos componentes de ganho durante a gestação, estimadas
através das equações propostas pelo NRC (2012).
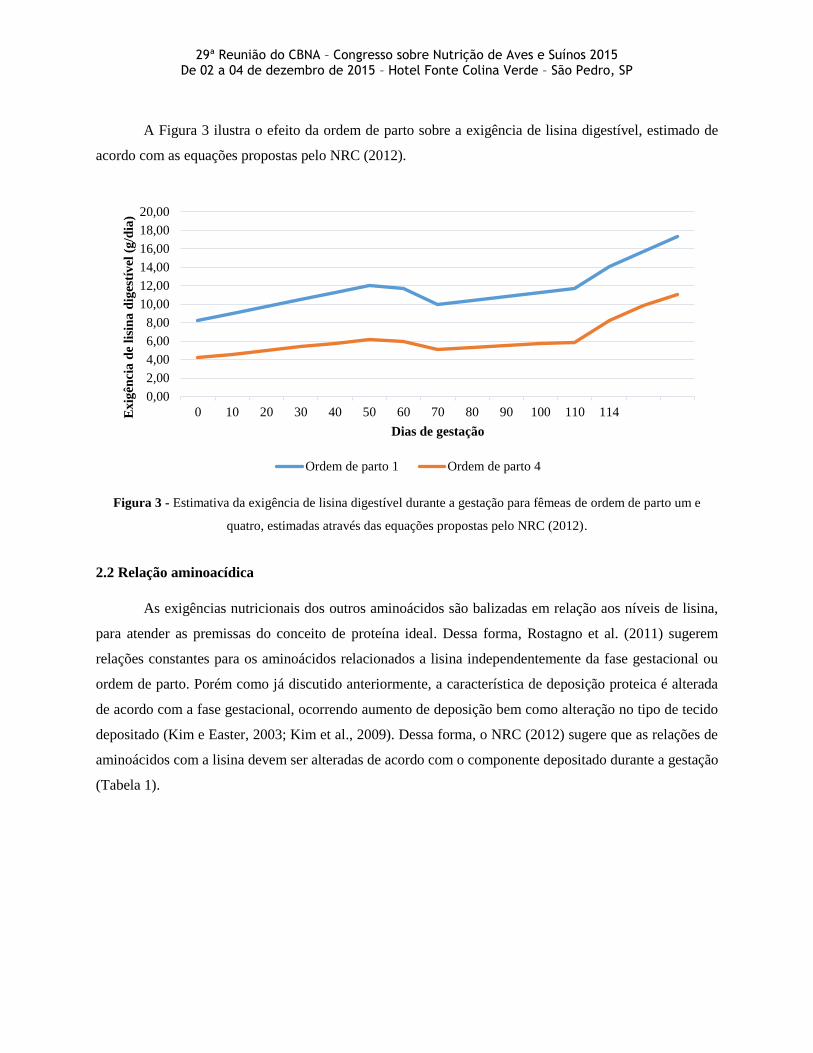
Na Figura 2 é demonstrado o efeito do tamanho da leitegada sobre a exigência de lisina digestível.
Neste sentido, faz-se necessário reconhecer o histórico da granja para adaptar a dieta de acordo com a
capacidade produtiva das matrizes suínas.
Figura 2 - Estimativa da exigência de lisina digestível durante a gestação para fêmeas com leitegadas de 14 ou 12
leitões, estimadas através das equações propostas pelo NRC (2012).
0
20
40
60
80
100
120
140
0 10 20 30 40 50 60 70 80 90 100 110 114
Dep
osi
ção p
rote
ica (
g/d
ia)
Dias de gestação
Deposição proteica total Deposição proteica fetal
Deposição proteica mamária Deposição proteica placenta e fluídos
Deposição proteica uterina
1,00
3,00
5,00
7,00
9,00
11,00
0 10 20 30 40 50 60 70 80 90 100 110 114Exig
ênci
a d
e li
sin
a d
iges
tív
el (
g/d
ia)
Dias de gestação
Total de 16 leitões Total de 14 leitões
29ª Reunião do CBNA – Congresso sobre Nutrição de Aves e Suínos 2015 De 02 a 04 de dezembro de 2015 – Hotel Fonte Colina Verde – São Pedro, SP
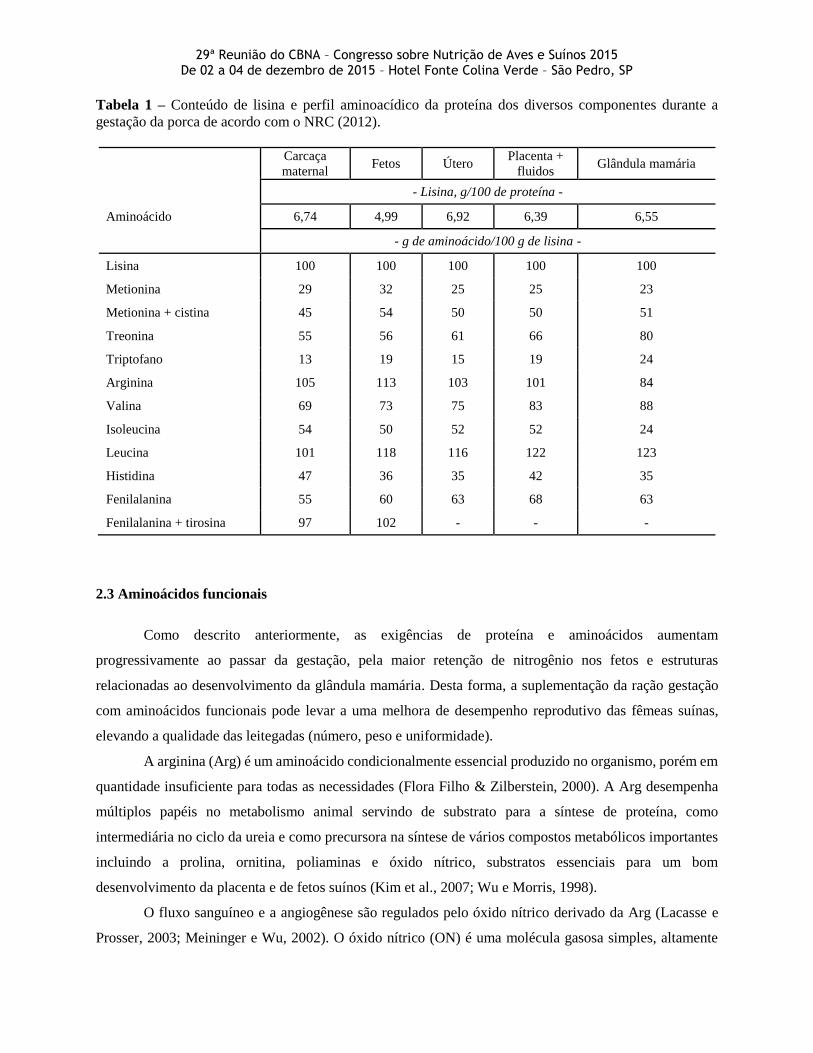
A Figura 3 ilustra o efeito da ordem de parto sobre a exigência de lisina digestível, estimado de
acordo com as equações propostas pelo NRC (2012).
Figura 3 - Estimativa da exigência de lisina digestível durante a gestação para fêmeas de ordem de parto um e
quatro, estimadas através das equações propostas pelo NRC (2012).
2.2 Relação aminoacídica
As exigências nutricionais dos outros aminoácidos são balizadas em relação aos níveis de lisina,
para atender as premissas do conceito de proteína ideal. Dessa forma, Rostagno et al. (2011) sugerem
relações constantes para os aminoácidos relacionados a lisina independentemente da fase gestacional ou
ordem de parto. Porém como já discutido anteriormente, a característica de deposição proteica é alterada
de acordo com a fase gestacional, ocorrendo aumento de deposição bem como alteração no tipo de tecido
depositado (Kim e Easter, 2003; Kim et al., 2009). Dessa forma, o NRC (2012) sugere que as relações de
aminoácidos com a lisina devem ser alteradas de acordo com o componente depositado durante a gestação
(Tabela 1).
0,00
2,00
4,00
6,00
8,00
10,00
12,00
14,00
16,00
18,00
20,00
0 10 20 30 40 50 60 70 80 90 100 110 114Exig
ênci
a d
e li
sin
a d
iges
tív
el (
g/d
ia)
Dias de gestação
Ordem de parto 1 Ordem de parto 4
29ª Reunião do CBNA – Congresso sobre Nutrição de Aves e Suínos 2015 De 02 a 04 de dezembro de 2015 – Hotel Fonte Colina Verde – São Pedro, SP
Tabela 1 – Conteúdo de lisina e perfil aminoacídico da proteína dos diversos componentes durante a
gestação da porca de acordo com o NRC (2012).
Carcaça
maternal Fetos Útero
Placenta +
fluidos Glândula mamária
- Lisina, g/100 de proteína -
Aminoácido 6,74 4,99 6,92 6,39 6,55
- g de aminoácido/100 g de lisina -
Lisina 100 100 100 100 100
Metionina 29 32 25 25 23
Metionina + cistina 45 54 50 50 51
Treonina 55 56 61 66 80
Triptofano 13 19 15 19 24
Arginina 105 113 103 101 84
Valina 69 73 75 83 88
Isoleucina 54 50 52 52 24
Leucina 101 118 116 122 123
Histidina 47 36 35 42 35
Fenilalanina 55 60 63 68 63
Fenilalanina + tirosina 97 102 - - -
2.3 Aminoácidos funcionais
Como descrito anteriormente, as exigências de proteína e aminoácidos aumentam
progressivamente ao passar da gestação, pela maior retenção de nitrogênio nos fetos e estruturas
relacionadas ao desenvolvimento da glândula mamária. Desta forma, a suplementação da ração gestação
com aminoácidos funcionais pode levar a uma melhora de desempenho reprodutivo das fêmeas suínas,
elevando a qualidade das leitegadas (número, peso e uniformidade).
A arginina (Arg) é um aminoácido condicionalmente essencial produzido no organismo, porém em
quantidade insuficiente para todas as necessidades (Flora Filho & Zilberstein, 2000). A Arg desempenha
múltiplos papéis no metabolismo animal servindo de substrato para a síntese de proteína, como
intermediária no ciclo da ureia e como precursora na síntese de vários compostos metabólicos importantes
incluindo a prolina, ornitina, poliaminas e óxido nítrico, substratos essenciais para um bom
desenvolvimento da placenta e de fetos suínos (Kim et al., 2007; Wu e Morris, 1998).
O fluxo sanguíneo e a angiogênese são regulados pelo óxido nítrico derivado da Arg (Lacasse e
Prosser, 2003; Meininger e Wu, 2002). O óxido nítrico (ON) é uma molécula gasosa simples, altamente

29ª Reunião do CBNA – Congresso sobre Nutrição de Aves e Suínos 2015 De 02 a 04 de dezembro de 2015 – Hotel Fonte Colina Verde – São Pedro, SP
lipofílica e é o maior vasodilatador das células endoteliais (Wu e Meininger, 2000), desempenhando um
papel importante na regulação do fluxo sanguíneo placentário e, portanto, na transferência de nutrientes e
oxigênio da mãe para o feto (Bird et al., 2003). Além disso, o ON é um importante mensageiro intercelular
nos mamíferos superiores. O mecanismo de sinalização intercelular é, em geral, realizado através de
receptores de membrana celular na célula alvo e, habitualmente, são transmembranosos em contato com
citoplasma e desencadeando uma “cascata” de sinais intracelulares que interfere no metabolismo celular.
A ativação da produção de poliaminas pela Arg, tem papel importante, já que esta molécula regula
o DNA e a síntese proteica, estando diretamente relacionadas à proliferação e diferenciação celular e ainda
contribui através da via sinalizadora do mTOR (Wu et al., 2011).
São vários os benefícios relacionados a utilização de Arg para fêmeas em gestação, entre eles pode-
se citar:
1) Aumento em 9,2% do número total de leitões nascidos (Wu et al., 2010; Gao et al., 2012);
2) Aumento do número de leitões nascidos vivos em 12,0% (Wu et al., 2010; Gao et al., 2012);
3) Diminuição de leitões natimortos em 59,8% (Mateo et al., 2007; Wu et al., 2010; Liu et al., 2012);
4) Aumento do peso ao nascer dos leitões desmamados em 3,9% (Liu et al., 2012; Garbossa et al., 2015);
5) Aumento do número de fetos viáveis em 30,4% (Bérard e Bee, 2010; Li et al., 2014).
Já a glutamina (Glu) apresenta concentrações elevadas no tecido proteico dos fetos, sendo que a
sua deficiência é o principal fator que contribui para o aparecimento de crescimento intrauterino retardado
(CIUR) (Wu et al., 2011). Da mesma forma que a Arg, a Glu ativa a síntese proteica através da produção
de poliaminas e ativação do mTOR. Neste sentido, o fornecimento deste aminoácido na dieta de matrizes
em gestação traz vantagens como aumento do peso do leitão ao nascimento e redução de CIUR.
Wu et al. (2010) avaliando a associação de Arg e Glu para fêmeas suínas verificou o aumento do
peso ao nascimento da leitegada em 9,6%, e diminuição da quantidade de leitões natimortos em 49,6%
quando comparados ao grupo controle. Da mesma forma, Wu et al. (2011) fornecendo Glu para as fêmeas
em gestação verificaram aumento do peso ao nascimento dos leitões em 6,0%.
3. Lactação
3.1 Dinâmica dos aminoácidos durante a lactação
Para o período de lactação, a taxa de crescimento da leitegada é um fator determinante na escolha
de programas nutricionais. O objetivo de minimizar possíveis impactos negativos nos ciclos reprodutivos
seguintes também deve ser considerado. As fêmeas suínas contemporâneas foram selecionadas para uma

29ª Reunião do CBNA – Congresso sobre Nutrição de Aves e Suínos 2015 De 02 a 04 de dezembro de 2015 – Hotel Fonte Colina Verde – São Pedro, SP
alta prolificidade e produção de leite, gerando assim uma alta exigência nutricional. Em contrapartida,
possuem alta capacidade de deposição de tecido magro, baixas reservas lipídicas e baixo consumo
voluntário (Fontes et al., 2014).
As exigências nutricionais, e principalmente de aminoácidos, podem variar bastante entre as
fêmeas e serem dependentes de fatores como peso corporal, composição corporal, genética, idade,
capacidade de produção de leite, entre outros. Condições de ambiente e de alimentação também podem ser
consideradas fontes de variação, sendo que estas, diferentemente dos fatores individuais, podem ser
controladas (Strathe et al., 2015). Conhecer tais fatores é essencial para otimizar a produtividade e
proporcionar longevidade a fêmea. Além disso, torna-se uma ferramenta valiosa para o desenvolvimento
de estratégias nutricionais.
A fêmea lactante moderna é incapaz de ingerir o suficiente para satisfazer sua alta necessidade
nutricional (Bergsma et al., 2009; Quiniou, 2015) e este é o principal desafio do nutricionista em relação
ao plantel reprodutivo. Menos de 18% das fêmeas apresentam o padrão ideal de consumo alimentar na
lactação (Koketsu et al., 1996), afetando diretamente o crescimento da glândula mamária e a produção de
leite (Kim et al, 1999; De Bettio et al., 2015). Além disso, uma mobilização proteica excessiva geralmente
resulta em falhas reprodutivas subsequentes (Vinsky et al., 2006; De Bettio et al., 2015).
Para pluríparas, esta perda pode ser de no máximo 9 a 12% da massa proteica, não
comprometendo assim seu desempenho reprodutivo posterior (Clowes et al. 2003). Já em primíparas, a taxa
de mobilização não deve chegar a 8% do seu peso corporal (Schenkel et al., 2010).
A exigência de lisina em porcas lactantes pode ser estimada pelo modelo fatorial a partir da
diferença entre as exigências para mantença e para produção de leite e a quantidade fornecida por meio da
mobilização da proteína corporal. Para se determinar as exigências nutricionais de fêmeas suínas lactantes,
Rostagno et al. (2011) utilizam como parâmetros o peso corporal, o ganho de peso da leitegada e a perda
de peso da fêmea durante a lactação. Já o NRC (2012) utiliza o tamanho da leitegada, o ganho de peso dos
leitões e a produção de leite. Para Dourmad et al. (2008), avaliar a eficiência de utilização de aminoácidos
dietéticos para a produção de leite é difícil, já que a contribuição das reservas proteicas maternas não é
conhecida com precisão. A quantidade e a composição das reservas corporais mobilizadas durante a
lactação irão depender do déficit nutricional da fêmea.
Os critérios utilizados para definir as exigências de aminoácidos são determinantes, já que as
necessidades são diferentes para o crescimento da leitegada e para a mínima mobilização de tecido corporal
materno. Alguns autores confirmam que a exigência diária de lisina, para fêmeas primíparas, objetivando
minimizar a perda de peso corporal e não gerar falhas reprodutivas no ciclo seguinte é mais alta do que a
exigência para produção de leite (Tokach et al, 1999; Kim et al., 2001; Paiva et al., 2005). As porcas
multíparas não possuem necessidades nutricionais significativas de crescimento e têm uma capacidade

29ª Reunião do CBNA – Congresso sobre Nutrição de Aves e Suínos 2015 De 02 a 04 de dezembro de 2015 – Hotel Fonte Colina Verde – São Pedro, SP
relativamente maior de consumo de ração, além das reservas proteica e lipídica mais elevadas, o que resulta
em um estado catabólico menos dramático. Por isso, a porcentagem de lisina, em cada quilo de ração
fornecida para marrãs, deve ser maior do que a de fêmeas multíparas (Yan et al., 2009).
As diferenças nas recomendações para ingestão de lisina são comuns, seja entre tabelas nacionais
e estrangeiras, como entre as próprias empresas de genética. A exigência de lisina para Rostagno et al.
(2011) e NRC (2012) chega a diferenciar-se em 25% para classes de mesmo peso. Para as fêmeas mais
pesadas essa disparidade diminui (Tabela 2).
Tabela 2 – Exigência de lisina, em gramas por dia, em função do peso de fêmeas lactantes*.
Peso (kg) 175 210
Rostagno et al. (2011) (g/dia) 57,23 57,49
NRC (201) (g/dia) 45,70 48,90
Variação (%) 25,23 17,56
*2,5 kg/dia de ganho de peso da leitegada e 0,5 kg de perda de peso da fêmea.
Segundo Strathe et al. (2015), as exigências diárias de proteína e aminoácidos mudam ao longo
da lactação, sendo mais baixas na primeira semana de lactação, intermediárias na segunda e mais altas na
última semana. Mesmo com menor exigência aminoacídica na primeira semana, o ideal é trabalhar uma
dieta com maior relação lisina/energia, já que o consumo neste período é reduzido.
3.2 Relação aminoacídica
O conceito de proteína ideal significa a redução dos níveis de proteína bruta com o objetivo de
atender à exigência de aminoácidos essenciais com exatidão, respeitando a relação destes com a lisina,
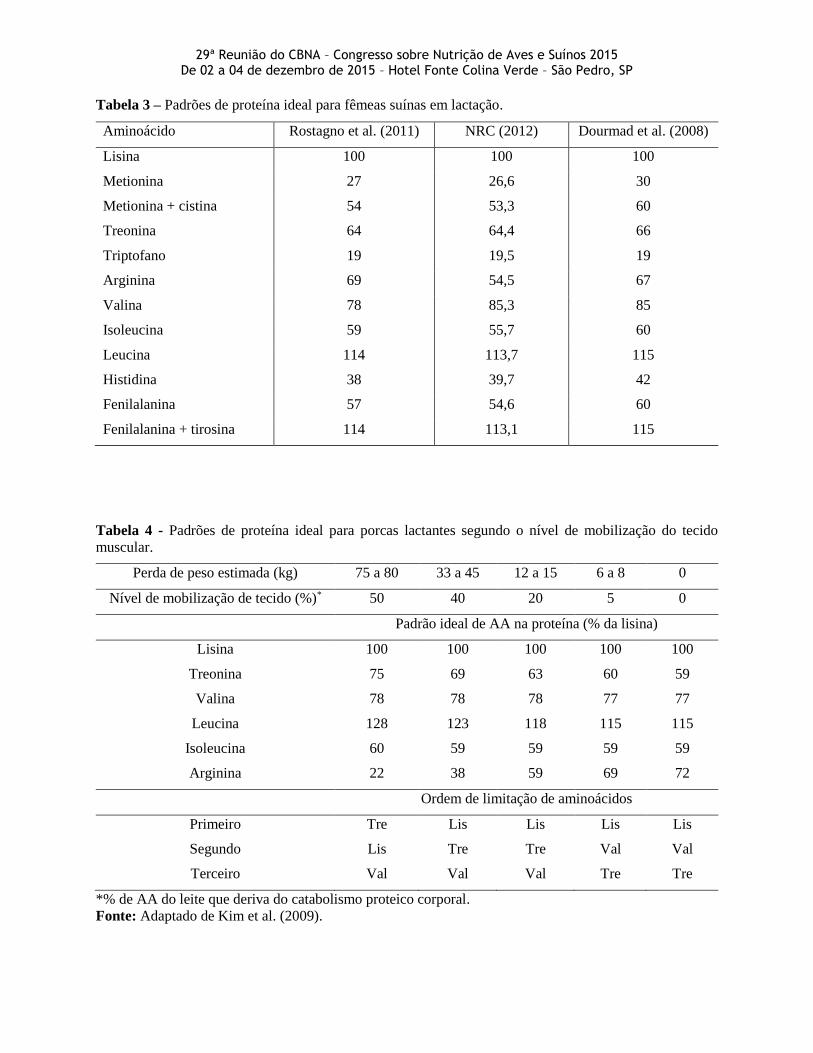
reduzindo os custos e a excreção de proteína. A Tabela 3 demonstra as diferenças entre as recomendações,
sendo que a arginina é o aminoácido que apresenta maior variação entre as fontes.
Segundo Kim et al (2001), a relação ideal entre os aminoácidos pode variar caso a diferença entre
os aminoácidos utilizados na produção de leite e os mobilizados pela fêmea se altere. O conceito de proteína
ideal dinâmico permite uma estimativa mais precisa das necessidades de porcas lactantes. A lisina é o
primeiro aminoácido limitante para fêmeas de variados consumos voluntários e taxas de mobilização. Já
para porcas de baixo consumo voluntário e, consequentemente, alta mobilização, a ingestão de treonina
torna-se cada vez mais importante.
29ª Reunião do CBNA – Congresso sobre Nutrição de Aves e Suínos 2015 De 02 a 04 de dezembro de 2015 – Hotel Fonte Colina Verde – São Pedro, SP
Tabela 3 – Padrões de proteína ideal para fêmeas suínas em lactação.
Aminoácido Rostagno et al. (2011) NRC (2012) Dourmad et al. (2008)
Lisina 100 100 100
Metionina 27 26,6 30
Metionina + cistina 54 53,3 60
Treonina 64 64,4 66
Triptofano 19 19,5 19
Arginina 69 54,5 67
Valina 78 85,3 85
Isoleucina 59 55,7 60
Leucina 114 113,7 115
Histidina 38 39,7 42
Fenilalanina 57 54,6 60
Fenilalanina + tirosina 114 113,1 115
Tabela 4 - Padrões de proteína ideal para porcas lactantes segundo o nível de mobilização do tecido
muscular.
Perda de peso estimada (kg) 75 a 80 33 a 45 12 a 15 6 a 8 0
Nível de mobilização de tecido (%)* 50 40 20 5 0
Padrão ideal de AA na proteína (% da lisina)
Lisina 100 100 100 100 100
Treonina 75 69 63 60 59
Valina 78 78 78 77 77
Leucina 128 123 118 115 115
Isoleucina 60 59 59 59 59
Arginina 22 38 59 69 72
Ordem de limitação de aminoácidos
Primeiro Tre Lis Lis Lis Lis
Segundo Lis Tre Tre Val Val
Terceiro Val Val Val Tre Tre
*% de AA do leite que deriva do catabolismo proteico corporal.
Fonte: Adaptado de Kim et al. (2009).

29ª Reunião do CBNA – Congresso sobre Nutrição de Aves e Suínos 2015 De 02 a 04 de dezembro de 2015 – Hotel Fonte Colina Verde – São Pedro, SP
3.3 Aminoácidos funcionais
O uso de aminoácidos funcionais já é uma realidade na produção suinícola e diversos trabalhos
demonstram os benefícios gerados pela inclusão desses aminoácidos na dieta de lactação.
O leite materno é deficiente em arginina (Wu et al., 2004), o que pode comprometer o
desenvolvimento dos leitões lactentes. Estudos recentes demonstram que a suplementação com L-arginina
na ração de lactação melhora o ganho de peso dos leitões, já que existem um enriquecimento na qualidade
nutricional do leite (Mateo et al, 2008; Moreira, 2014). A arginina também parece aumentar a eficiência da
utilização da proteína dietética para a síntese do leite, através da melhora no fluxo sanguíneo e no
fornecimento de nutrientes para a glândula mamária (Mateo et al., 2008).
Já a glutamina é o aminoácido livre mais abundante no leite materno. Mas, uma vez que este
aminoácido é metabolizado pelas células epiteliais do intestino delgado, há pouca absorção líquida em
níveis normais de consumo alimentar. Por isso, a suplementação de fêmeas com este aminoácido funcional
se torna bastante interessante. Marrãs lactantes suplementadas com L-glutamina apresentam menor perda
de massa corporal magra e maior concentração de glutamina no leite (Manso et al., 2012). Este aumento
pode ser acompanhado pelo aumento no teor de gordura do leite, desde o colostro até o final da lactação
(Santos de Aquino, 2014). Tal abordagem oferece um mecanismo muito atraente para aumentar a
disponibilidade de glutamina para o leitão lactente, já que está relacionada não só com situações de extremo
catabolismo, mas com a manutenção da saúde e maiores taxas de crescimento (Watford, 2015).
4. Conclusão
A quantidade exata de nutrientes exigidos, fornecida de forma correta ao animal, é o principal
objetivo da nutrição de precisão. Para o plantel reprodutivo, os ajustes individuais e dinâmicos são ainda
mais importantes. Além disto, as matrizes suínas modernas são desafiadas a atingirem índices produtivos
cada vez mais altos, gerando um grande desgaste metabólico e uma alta demanda nutricional.
Desta maneira, modelos matemáticos estão sendo desenvolvidos com o objetivo de entender e
atender os requisitos de uma população de fêmeas que se diferem em peso, composição corporal, tamanho
de leitegada e produção de leite.
Os programas de alimentação para matrizes gestantes devem ser revistos no intuito de ofertar uma
maior quantidade de aminoácidos na segunda metade da gestação, apoiando assim, o aumento da utilização
de proteínas para o crescimento da glândula mamária e dos fetos. Se a estratégia é adotada e um padrão
adequado de aminoácidos é fornecido, minimiza-se o ganho de gordura desnecessário durante este período
gestacional. Por isso, duas dietas na gestação são cada vez mais usadas.
Fêmeas lactantes requerem um ajuste preciso na dieta de acordo com o desempenho, mas também
às condições de alojamento, o que pode afetar a utilização de nutrientes e a ingestão voluntária dos animais.

29ª Reunião do CBNA – Congresso sobre Nutrição de Aves e Suínos 2015 De 02 a 04 de dezembro de 2015 – Hotel Fonte Colina Verde – São Pedro, SP
Condições ambientais também interferem diretamente na ingestão de ração e no direcionamento dos
nutrientes.
Sendo assim, ferramentas de individualização do arraçoamento serão cada vez mais úteis quando
pensamos em uma nutrição precisa e com excelência.
5. Referências bibliográficas
BÉRARD, J. E.; BEE, G. Effects of dietary l-arginine supplementation to gilts during early gestation on
foetal survival, growth and myofiber formation. Animal, v. 4, p. 1680-1687, 2010.
BERGSMA, R. et al. Lactation efficiency as a result of body composition dynamics and feed intake in
sows. Livestock Science, v. 125, p. 208-222, 2009.
BIRD, I. M.; ZHANG, L. B.; MAGNESS, R. R. Possible mechanisms underlying pregnancyinduced
changes in uterine artery endothelial function. American Journal of Physiology, Baltimore, v. 284, p.
R245-R258, 2003.
CARPENTER, K. J. A short history of nutritional science: Part 1 (1785–1885). Journal of Nutrition,
v.133, p. 638–645, 2003a.
CARPENTER, K. J. A short history of nutritional science: Part 2 (1885–1912). Journal of Nutrition,
v.133, p. 975–984, 2003b.
CLOSE, W. H.; MULLAN, B. P. Nutrition and feeding of breeding stock. In: TAVORNER, M.R.,
DUNKIN, A.C., Pig Production, Elsevier, New York, 1996.
CLOWES, E. J. et al. Parturition body size and body protein loss during lactation influence performance
during lactation and ovarian function at weaning in first-parity sow. Journal of Animal Science, v. 81, p.
1517-1528, 2003.
DE BETTIO, S. et al. Impact of feed restriction on the performance of highly prolific lactating sows and
its effect on the subsequent lactation. Animal, p. 1-7, 2015.
DOURMAD, J. et al. InraPorc: A model and decision support tool for the nutrition of sows. Animal Feed
Science and Tecnology, v. 143, p. 372-386, 2008.
FLORA FILHO, R.; ZILBERSTEIN, B. Óxido nítrico: o simples mensageiro percorrendo a
complexidade. Metabolismo, síntese e funções. Revista da Associação Medica Brasileira, São Paulo, v.
46, n. 3, p. 265-271, 2000.
FONTES, D. O.; ABREU, M. L. T; NETA, C. S. S. Exigências nutricionais da fêmea suína lactante. In:
Associação Brasileira de Criadores de Suínos. Produção de suínos: teoria e prática. Brasília:
Associação Brasileira de Criadores de Suínos, 2014, p. 507-516.
GAO, K. et al. Dietary L -arginine supplementation enhances placental growth and reproductive
performance in sows. Amino Acids, v. 42, p. 2207-2214, 2012.
GARBOSSA. C. A. P. et al. Effects of ractopamine and arginine dietary supplementation for sows on
growth performance and carcass quality of their progenies. Journal of Animal Science, v. 6, p. 2872-
2884, 2015.

29ª Reunião do CBNA – Congresso sobre Nutrição de Aves e Suínos 2015 De 02 a 04 de dezembro de 2015 – Hotel Fonte Colina Verde – São Pedro, SP
HUANG, F. R. et al. Effects of lysine and protein intake over two consecutive lactations on lactation and
subsequent reproductive performance in multiparous sows. Livestock Science, v.157, p. 482–489, 2013.
JI , F., HURLEY, W. L., KIM, S. W. Characterization of mammary gland development in pregnant gilts.
Journal of Animal Science, v. 84, p. 579-587, 2006.
JONES, D. B.; STAHLY,T. S. Impact of amino acid nutrition during lactation on body nutrient
mobilization and milk nutrient output in primiparous sows. Journal of Animal Science, v. 77, p. 1513–
1522, 1999.
KIM, S.W. et al. Effect of nutrient intake on mammary gland growth in lactating sows. Journal of
Animal Science, v. 77, p. 3304-3315, 1999.
KIM, S.W . BAKER, D. H.; EASTER, R. A. Dynamic ideal protein and limiting amino acids for lactating
sows: The impact of amino acid mobilization. Journal of Animal Science, v. 79, p. 2356-2366, 2001.
KIM, S. W.; EASTER, R. A. Amino acid utilization for reproduction in sows. In: D’MELLO, J. P. F.
Amino acids in animal nutrition. 2nd ed. Edinburgh, UK: CAB Publishing, 2003. p. 203-222.
KIM, S. W. et al. Ideal amino acid balance for sows during gestation and lactation. Journal of Animal
Science, v. 87, p 123-132, 2009.
KIM, S. W. et al. Functional amino acids and fatty acids for enhancing production performance of sows
and piglets. Asian-Australasian Journal of Animal Sciences, Seoul, v. 20, p. 295–306, 2007.
KOKETSU, Y. et al. Feed intake pattern during lactation and subsequent reproductive performance of
sows. Journal of Animal Science, v. 74, n. 12, p. 2875-2884, 1996.
LACASSE, P.; PROSSER, C. G. Mammary blood flow does not limit milk yield in lactating goats.
Journal of Dairy Science, Champaign, v. 86, p. 2094–2097, 2003.
LI, X. et al. Dietary supplementation with L-arginine between days 14 and 25 of gestation enhances
embryonic development and survival in gilts. Amino Acids, v. 46, p. 375-384, 2014.
LIU, X. et al. Effects of dietary L-arginine or N-carbamylglutamate supplementation during late gestation
of sows on the miR-15b/16, miR-221/222, VEGFA and eNOS expression in umbilical vein. Amino
Acids, v. 42, p. 2111-2119, 2012.
MANSO, H. E. C. C. C. et al. Glutamine and glutamate supplementation raise milk glutamine
concentrations in lactating gilts. Journal of Animal Science and Biotechnology, v. 3, p 1-7, 2012. MATEO, R. D. et al. Effects of dietary arginine supplementation during gestation and lactation on the
performance of lactating primiparous sows and nursing piglets. Journal Animal Science, v.86, n.4,
p.827-835, 2008.
MATEO, R. D. et al. Dietary l-arginine supplementation enhances the reproductive performance of gilts.
Journal of Nutrition, v. 137, p. 652-656. 2007.
MCPHERSON, R. L. et al. Growth and compositional changes of fetal tissues in pigs. Journal of Animal
Science, v. 82, p. 2534-2540, 2004.
MEININGER, C. J.; WU, G. Regulation of endothelial cell proliferation by nitric oxide. Methods in
Enzymology, New York, v. 352, p. 280–295, 2002.

29ª Reunião do CBNA – Congresso sobre Nutrição de Aves e Suínos 2015 De 02 a 04 de dezembro de 2015 – Hotel Fonte Colina Verde – São Pedro, SP
MOEHN ,S., BALL, R.O. London swine conference. Managing for Production. 2013. p. 55–63. 3-27-
2013.
MOREIRA, R.H.R. Arginina na nutrição de matrizes suínas hiperprolíficas. 2014. 50 p. Dissertação
(Mestrado em Zootecnia). Universidade Federal de Lavras, Lavras, 2014.
MUNRO, H.N., ALLISON, J.B., editors. Mammalian Protein Metabolism, Vol II. New York:
Academic Press; 1964.
NATIONAL RESEARCH COUNCIL. Nutrient Requirements of Swine. 11. ed. Washington, D. C.:
National Academy, 2012. 400 p.
PAIVA, F. P. et al. Lisina em rações para fêmeas suínas primíparas em lactação. Revista Brasileira de
Zootecnia, Viçosa, MG, v. 34, n. 6, p. 1971-1979, 2005.
QUINIOU, N. Amino acid nutrition of the prolific sow. In: Congresso ABRAVES, 17., 2015, Campinas.
Anais… Campinas: Embrapa Aves e Suínos, 2015. p. 28-33.
ROSTAGNO, H.S. et al. Tabelas brasileiras para aves e suínos: composição de alimentos e exigências
nutricionais. 3. ed. Viçosa: UFV, 2011. 252 p.
SANTOS DE AQUINO, R. et al. Glutamine and glutamate (AminoGut) supplementation influences
sow colostrum and mature milk composition. Livestock Science, v. 169, 112-117, 2014.
SCHENKEL , A.C. et al. Body reserve mobilization during lactation in first parity sows and its effect on
second litter size. Livestock Science, v. 132, p. 165-172, 2010.
STRATHE, A.V. et al. Determination of protein and amino acid requirements of lactating sows using a
population-based factorial approach. Animal, v. 9, p. 1319-1328, 2015.
TOKACH, M. et al. Nutrition for optimal performance of the female pig. In: PIG FARMERS’
CONFERENCE, 1999, [S.l]. Proceddings... [Carlow, Ireland: Teagasc], 1999. Disponível em:
<http://www.teagasc.ie/publications/pig1999/paper12.asp>. Acesso em: 22 apr. 2004;
VINSKY, M.D. et al. Nutritional restriction in lactating primiparous sows selectively affects female
embryo survival and overall litter development. Reproduction Fertility Development, v. 18, p. 347–
355, 2006.
WATFORD, M. Glutamine and glutamate: Nonessential or essential amino acids? Animal Nutrition,
2015, http://dx.doi.org/10.1016/j.aninu.2015.08.008.
WU, G. et al. Triennial growth symposium: Important roles for l-glutamine in swine nutrition and
production. Journal of Animal Science, v. 89, p. 2017-2030, 2011.
WU, G.; KNABE, D.A.; KIM, S.W. Arginine nutrition in neonatal pigs. Journal of Nutrition, v. 134, n.
10, p. 2783-2790, 2004.
WU, G.; MEININGER, C. J. Arginine nutrition and cardiovascular function. Journal of Nutrition, v.
130, p. 2626–2629, 2000.
WU, G.; MORRIS JÚNIOR, S. M. Arginine metabolism: nitric oxide and beyond. Biochemical Journal,
London, v. 336, p. 1-17, 1998.

29ª Reunião do CBNA – Congresso sobre Nutrição de Aves e Suínos 2015 De 02 a 04 de dezembro de 2015 – Hotel Fonte Colina Verde – São Pedro, SP
WU, X. et al. Dietary supplementation with l-arginine or N-carbamylglutamate enhances intestinal
growth and heat shock protein-70 expression in weanling pigs fed a corn- and soybean meal-based diet.
Amino Acids, 39, 831-839, 2010.
YAN, Y.X. et al. Effects of lysine intake during late gestation and lactation on blood metabolites,
hormones, milk composition and reproductive performance in primiparous and multiparous sows. Animal
Reproduction Science, v. 112, p. 199-214, 2009.


















