
Allo-class i-reactive cd4-cd8- t cell hybridomas recognize the conformational change of class i...
5
Eur. J. Immunol. 1989. 19: 1861-1865 The role of a3 domain of class I molecule 1861 Junji Yamamoto, Manabu Tanabe, Mutsuhiko MinamiO, Kyoichi Kano and Masafumi Takiguchi Department of Immunology, Institute of Medical Science and Department of Blood Transfusiono, School of Medicine, University of Tokyo, Tokyo Allo-class I-reactive CD4-CDtt T cell hybridomas recognize the conformational change of class I molecule resulting from the exchange of the a3 domain* H-2Kb-reactive, interleukin (IL) 2-producingTcel1 hybridomas possessing neither CD8 nor CD4 molecules were employed for the study of allorecognition on interspecies hybrid antigens by T cells in the absence of an influence of these accessory molecules. Both HTB176.10 and HTB177.2 Tcell hybridomas reacted with KbKbB7 hybrid antigens as well as Kb antigens and they produced more IL 2 in response to Kb antigens than KbKbB7 hybrid antigens. IL2 production of these Tcell hybridomas was dependent on the surface expression level of Kb molecules on stimulators. Therefore, L cells expressing almost equivalent levels of Kb or hybrid antigens were selected for further functional studies by these T cell hybridomas. They apparently produced less IL2 in response not only to interspecies hybrid antigens but also to interspecies hybrid antigens in response to Kb antigens. These results indicated that T cell hybridomas recognized the conformational change of class I molecule resulting from the exchange of the a3 domain via their T cell receptors. 1 Introduction MHC class I molecules are expressed on most somatic cells and play a central role in T cell recognition, including alloreactivity, anti-viral reaction and probably anti-tumor response. Functional analysis of hybrid class I molecule created by exon shuffling [l, 21 and of mutant class I molecules reacted by site-directed mutagenesis [3,4] demonstrated that a1 and a2 domains contained the alloantigenic determinants recognized by allospecific mAb and allospecific CTL. On the other hand, recent studies by Maziarz et al. [5] and Connolly et al. [6] demonstrated that the a3 domain of class I molecule also influenced to allorecognition by CTL. However, these studies using CTL could not completely determine whether failure of allo- recognition by CTL was due to the loss of binding site of the CD8 molecule or the change of allodeterminants by the exchange of the a3 domain. We have previously cloned six class I-reactive Tcell hybrid- omas and found that three of them recognized Kb antigens without co-recognition of class I1 antigens [7, 81. FCM analysis of these T cell hybridomas showed that they possessed neither CD8 and CD4 molecules. Therefore, we have chosen these hybridomas for studies of T cell recog- nition on interspecies and intraspecies hybrid class I anti- gens in the absence of any influence at the accessory molecules. Furthermore, our preliminary experiments had indicated that, in contrast to CTL clones, these T cell hybridomas could be utilized for quantitative analysis of T [I 75491 * Supported by Grant-in Aid for Scientific Research from the Ministry of Education, Culture and Science, the government of Japan. Correspondence: Masafumi Takiguchi, Department of Immunolo- gy, Institute of Medical Science, University of Tokyo, 4-6-1, Shirokanedai, Minato-ku, Tokyo 108, Japan cell recognition. In the present study, we have investigated allodeterminants of interspecies as well as intraspecies hybrid class I molecules recognized by the T cell hybrid- omas and found a role for the a3 domain in formation of the allodeterminants. 2 Materials and methods 2.1 Construction of hybrid genes and gene transfer into murine L cells Construction of hybrid genes between H-2Kb and HLA- B7, and their expression in L cells have been previously described [9]. Nomenclature of Kb/HLA-B7 hybrid genes was as follows: for example, KbKbB7 denotes a hybrid gene containing the a1 and a2 domains of Kb and the a3 domain of B7. For the construction of KbKbDp intraspecies hybrid genes, exons 1-3 of the Kbgene were linked to exons 4-8 of the DP gene: a 2.4-kb Bam HI fragment which contained exons 4-8 of the DP gene was isolated and then ligated into the Barn HI site of pUC18 containing the 1.9-kb Xba I frag- ment of exons 1-3 of the Kb gene. The hybrid gene was transfected into Ltk- cells by co-transfection with the HSV thymidine kinase gene using a modification of the calcium phosphate co-precipitation procedure. HAT-resistant co- lonies were selected and tested by indirect immunofluores- cence assay with anti-Kb mAb. 2.2 T cell hybridoma and mAb Two T cell hybridomas, HTB176.10 and HTB177.2, react with H-2Kb antigen and produce IL2 without co-recogni- tion of class I antigens [7, 81. Four anti-Kb mAb were used to detect Kb, KbKbDp and KbKbB7 antigens on transfected L cells. Hybridoma cells secreting the 34.1.2, B8.3.24 and 28.8.6 mAb were obtained from the American Type 0 VCH Verlagsgesellschaft mbH, D-6940 Weinheim, 1989 0014-2980/&9/1010-1861$02.50/0
-
Upload
junji-yamamoto -
Category
Documents
-
view
214 -
download
2
Transcript of Allo-class i-reactive cd4-cd8- t cell hybridomas recognize the conformational change of class i...

Eur. J. Immunol. 1989. 19: 1861-1865 The role of a3 domain of class I molecule 1861
Junji Yamamoto, Manabu Tanabe, Mutsuhiko MinamiO, Kyoichi Kano and Masafumi Takiguchi
Department of Immunology, Institute of Medical Science and Department of Blood Transfusiono, School of Medicine, University of Tokyo, Tokyo
Allo-class I-reactive CD4-CDtt T cell hybridomas recognize the conformational change of class I molecule resulting from the exchange of the a3 domain*
H-2Kb-reactive, interleukin (IL) 2-producingTcel1 hybridomas possessing neither CD8 nor CD4 molecules were employed for the study of allorecognition on interspecies hybrid antigens by T cells in the absence of an influence of these accessory molecules. Both HTB176.10 and HTB177.2 Tcell hybridomas reacted with KbKbB7 hybrid antigens as well as Kb antigens and they produced more IL 2 in response to Kb antigens than KbKbB7 hybrid antigens. IL2 production of these Tcell hybridomas was dependent on the surface expression level of Kb molecules on stimulators. Therefore, L cells expressing almost equivalent levels of Kb or hybrid antigens were selected for further functional studies by these T cell hybridomas. They apparently produced less IL2 in response not only to interspecies hybrid antigens but also to interspecies hybrid antigens in response to Kb antigens. These results indicated that T cell hybridomas recognized the conformational change of class I molecule resulting from the exchange of the a3 domain via their T cell receptors.
1 Introduction
MHC class I molecules are expressed on most somatic cells and play a central role in T cell recognition, including alloreactivity, anti-viral reaction and probably anti-tumor response. Functional analysis of hybrid class I molecule created by exon shuffling [l, 21 and of mutant class I molecules reacted by site-directed mutagenesis [3,4] demonstrated that a1 and a2 domains contained the alloantigenic determinants recognized by allospecific mAb and allospecific CTL. On the other hand, recent studies by Maziarz et al. [5] and Connolly et al. [6] demonstrated that the a3 domain of class I molecule also influenced to allorecognition by CTL. However, these studies using CTL could not completely determine whether failure of allo- recognition by CTL was due to the loss of binding site of the CD8 molecule or the change of allodeterminants by the exchange of the a3 domain.
We have previously cloned six class I-reactive Tcell hybrid- omas and found that three of them recognized Kb antigens without co-recognition of class I1 antigens [7, 81. FCM analysis of these T cell hybridomas showed that they possessed neither CD8 and CD4 molecules. Therefore, we have chosen these hybridomas for studies of T cell recog- nition on interspecies and intraspecies hybrid class I anti- gens in the absence of any influence at the accessory molecules. Furthermore, our preliminary experiments had indicated that, in contrast to CTL clones, these T cell hybridomas could be utilized for quantitative analysis of T
[I 75491
* Supported by Grant-in Aid for Scientific Research from the Ministry of Education, Culture and Science, the government of Japan.
Correspondence: Masafumi Takiguchi, Department of Immunolo- gy, Institute of Medical Science, University of Tokyo, 4-6-1, Shirokanedai, Minato-ku, Tokyo 108, Japan
cell recognition. In the present study, we have investigated allodeterminants of interspecies as well as intraspecies hybrid class I molecules recognized by the T cell hybrid- omas and found a role for the a3 domain in formation of the allodeterminants.
2 Materials and methods
2.1 Construction of hybrid genes and gene transfer into murine L cells
Construction of hybrid genes between H-2Kb and HLA- B7, and their expression in L cells have been previously described [9]. Nomenclature of Kb/HLA-B7 hybrid genes was as follows: for example, KbKbB7 denotes a hybrid gene containing the a1 and a2 domains of Kb and the a3 domain of B7.
For the construction of KbKbDp intraspecies hybrid genes, exons 1-3 of the Kb gene were linked to exons 4-8 of the DP gene: a 2.4-kb Bam HI fragment which contained exons 4-8 of the DP gene was isolated and then ligated into the Barn HI site of pUC18 containing the 1.9-kb Xba I frag- ment of exons 1-3 of the Kb gene. The hybrid gene was transfected into Ltk- cells by co-transfection with the HSV thymidine kinase gene using a modification of the calcium phosphate co-precipitation procedure. HAT-resistant co- lonies were selected and tested by indirect immunofluores- cence assay with anti-Kb mAb.
2.2 T cell hybridoma and mAb
Two T cell hybridomas, HTB176.10 and HTB177.2, react with H-2Kb antigen and produce IL2 without co-recogni- tion of class I antigens [7, 81. Four anti-Kb mAb were used to detect Kb, KbKbDp and KbKbB7 antigens on transfected L cells. Hybridoma cells secreting the 34.1.2, B8.3.24 and 28.8.6 mAb were obtained from the American Type
0 VCH Verlagsgesellschaft mbH, D-6940 Weinheim, 1989 0014-2980/&9/1010-1861$02.50/0
1862 J.Yamamoto, M. Tanabe, M. Minami et al.
Culture Collection (ATCC; Rockville, MD) and the 5041-16.3 mAb from Australian Monoclonal Development Pty Ltd. (Artamon, Australia).
Eur. J. Immunol. 1989. 19: 1861-1865
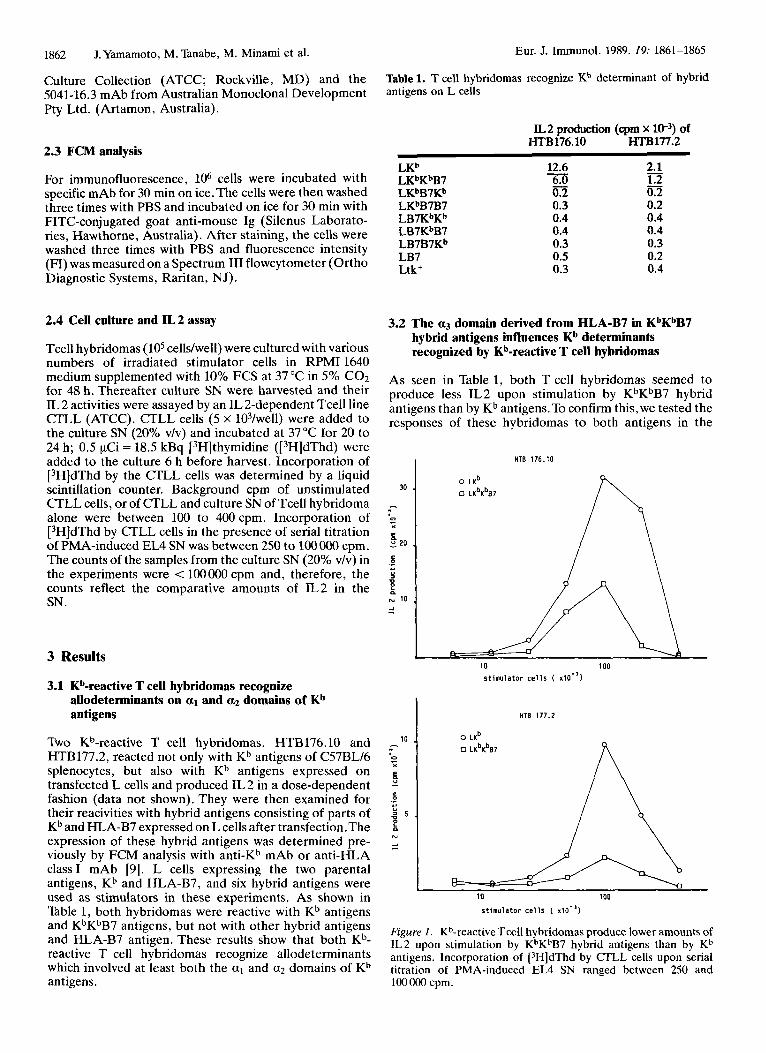
Table 1. T cell hybridomas recognize Kb determinant of hybrid antigens on L cells
2.3 FCM analysis
IL2 production (cpm x W3) of HTB176.10 HTBl77.2
12.6 2.1 For immunofluorescence, lo6 cells were incubated with L K ~ K ~ B ~ 6.0 1.2 specific mAb for 30 min on ice.The cells were then washed LKbB7Kb 0.2 0.2 three times with PBS and incubated on ice for 30 min with LKbB7B7 0.3 0.2
- LKb
FITC-conjugated goat anti-mouse Ig (Silenus Laborato- LB7KbKb 0.4 0.4 0.4 0.4 ries, Hawthorne, Australia). After staining, the cells were 0.3 0.3 0.5 0.2 washed three times with PBS and fluorescence intensity
Diagnostic Systems, Raritan, NJ).
LB7KbB7 LB7B7Kb LB7
(FI) was measured on a Spectrum I11 flowcytometer (Ortho Ltk+ 0.3 0.4
2.4 Cell culture and IL 2 assay
Tcell hybridomas (105 cells/well) were cultured with various numbers of irradiated stimulator cells in RPMI 1640 medium supplemented with 10% FCS at 37 "C in 5% C02 for 48 h. Thereafter culture SN were harvested and their IL 2 activities were assayed by an IL2-dependent Tcell line CTLL (ATCC). CTLL cells (5 x 103/well) were added to the culture SN (20% v/v) and incubated at 37°C for 20 to 24 h; 0.5 KCi = 18.5 kBq [3H]thymidine ([3H]dThd) were added to the culture 6 h before harvest. Incorporation of [3H]dThd by the CTLL cells was determined by a liquid scintillation counter. Background cpm of unstimulated CTLL cells, or of CTLL and culture SN of Tcell hybridoma alone were between 100 to 400cpm. Incorporation of [3H]dThd by CTLL cells in the presence of serial titration of PMA-induced EL4 SN was between 250 to 1oOOOO cpm. The counts of the samples from the culture SN (20% v/v) in the experiments were < 100000 cpm and, therefore, the counts reflect the comparative amounts of IL2 in the SN .
3 Results
3.1 Kb-reactive T cell hybridomas recognize allodeterminants on a1 and a2 domains of Kb antigens
Two Kb-reactive T cell hybridomas, HTB176.10 and HTB177.2, reacted not only with Kb antigens of C57BL/6 splenocytes, but also with Kb antigens expressed on transfected L cells and produced IL 2 in a dose-dependent fashion (data not shown). They were then examined for their reacivities with hybrid antigens consisting of parts of Kb and HLA-B7 expressed on L cells after transfection.The expression of these hybrid antigens was determined pre- viously by FCM analysis with anti-Kb mAb or anti-HLA class I mAb [9]. L cells expressing the two parental antigens, Kh and HLA-B7, and six hybrid antigens were used as stimulators in these experiments. As shown in Table 1, both hybridomas were reactive with Kb antigens and KbKbB7 antigens, but not with other hybrid antigens and HLA-B7 antigen. These results show that both Kh- reactive T cell hybridomas recognize allodeterminants which involved at least both the a1 and a2 domains of Kb antigens.
3.2 The a3 domain derived from HLA-B7 in KbKbB7 hybrid antigens influences Kb determinants recognized by Kb-reactive T cell hybridomas
As seen in Table 1, both T cell hybridomas seemed to produce less IL2 upon stimulation by KbKbB7 hybrid antigens than by Kb antigens.To confirm this, we tested the responses of these hybridomas to both antigens in the
HTE 176.10 I 30
- D L
I 0 20
c
u - Y e N 10 0
10
10 100 s t imula tor c e l l s ( x10-3 )
HTE 177.2
o L K ~ 0 LKbKbB7
10 100
s t imula tor ce l l s ( 110-3)
Figure 1. Kb-reactive Tcell hybridomas produce lower amounts of IL2 upon stimulation by KbKbB7 hybrid antigens than by Kb antigens. Incorporation of [3H]dThd by CTLL cells upon serial titration of PMA-induced EL4 SN ranged between 250 and 10OOOO cpm.
Eur. J. Immunol. 1989. 19: 1861-1865
Z , 0 L
280.
158
ie0
The role of a3 domain of class I molecule 1863
171.4
presence of different numbers of stimulator cells. As shown in Fig. 1, both hybridomas produced two-to threefold less IL 2 in response to KhKhB7 than in response to Kb at all cell doses tested. The lower production of IL2 by the hybrido- mas stimulated by KbKbB7 hybrid antigens was noted 24 h after the stimulation (data not shown).
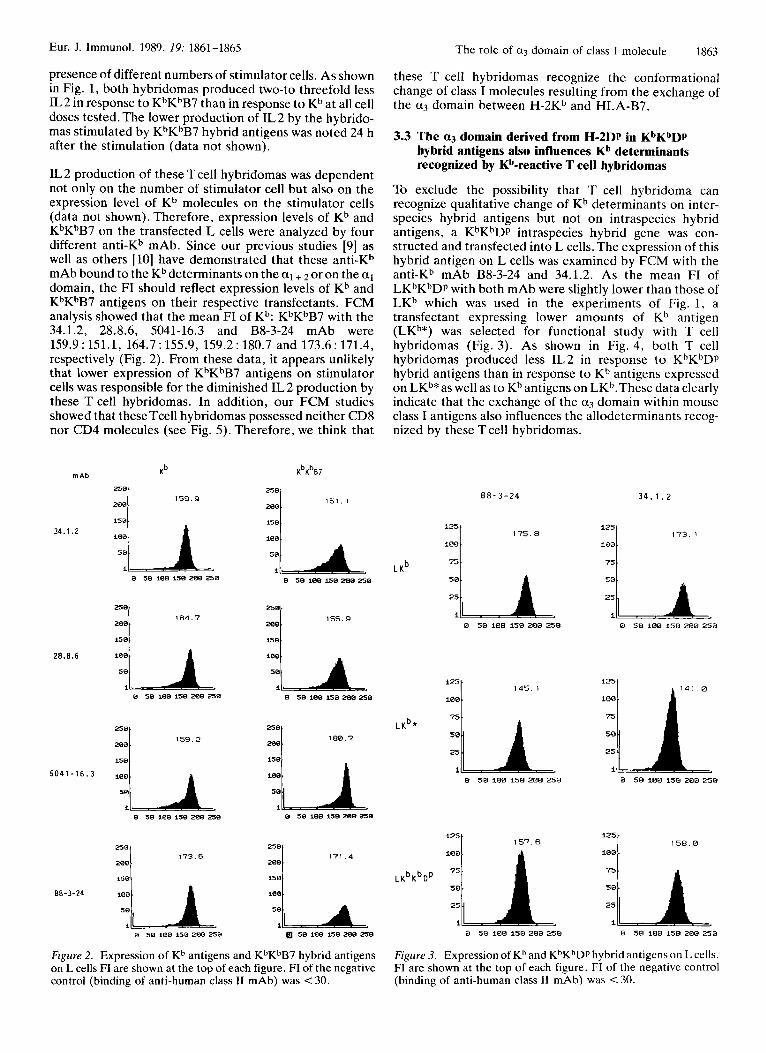
IL 2 production of these T cell hybridomas was dependent not only on the number of stimulator cell but also on the expression level of Kb molecules on the stimulator cells (data not shown). Therefore, expression levels of Kb and KbKbB7 on the transfected L cells were analyzed by four different anti-Kb mAb. Since our previous studies [9] as well as others [lo] have demonstrated that these anti-Kb mAb bound to the Kb determinants on the al + 2 or on the a1 domain, the FI should reflect expression levels of Kb and KbKbB7 antigens on their respective transfectants. FCM analysis showed that the mean FI of Kb: KbKhB7 with the 34.1.2, 28.8.6, 5041-16.3 and B8-3-24 mAb were 159.9 : 151.1, 164.7 : 155.9, 159.2 : 180.7 and 173.6 : 171.4, respectively (Fig. 2). From these data, it appears unlikely that lower expression of KbKbB7 antigens on stimulator cells was responsible for the diminished IL 2 production by these T cell hybridomas. In addition, our FCM studies showed that these Tcell hybridomas possessed neither CD8 nor CD4 molecules (see Fig. 5). Therefore, we think that
mAb
34.1 .2
28.8.6
5 0 4 1 -16.3
88-3-24
Kb
159.9
isal
I::/ . .
i
164.7
150
159.2
15w
0 50 I00 150 200 250
250 173.6
1581 ,1 . . I_ I
KbKb87
151. I
100 -at
We[ 155.9 280
0 50 100 158 280 258
180.7
15 8
50 100 158 200 250
Figure 2. Expression of Kh antigens and KhKhB7 hybrid antigens on L cells FI are shown at the top of each figure. FI of the negative control (binding of anti-human class I1 mAb) was < 30.
these T cell hybridomas recognize the conformational change of class I molecules resulting from the exchange of the a3 domain between H-2Kb and HLA-B7.
3.3 The a3 domain derived from H-2Dp in KbKbDp hybrid antigens also influences Kb determinants recognized by Kb-reactive T cell hybridomas
To exclude the possibility that T cell hybridoma can recognize qualitative change of Kb determinants on inter- species hybrid antigens but not on intraspecies hybrid antigens, a KhKhDp intraspecies hybrid gene was con- structed and transfected into L cells. The expression of this hybrid antigen on L cells was examined by FCM with the anti-Kb mAb B8-3-24 and 34.1.2. As the mean FI of LKbKhDp with both mAb were slightly lower than those of LKb which was used in the experiments of Fig. 1, a transfectant expressing lower amounts of Kb antigen (LKb*) was selected for functional study with T cell hybridomas (Fig. 3). As shown in Fig. 4, both T cell hybridomas produced less IL2 in response to KbKbDp hybrid antigens than in response to Kb antigens expressed on LKb*as well as to Kb antigens on LKb.These data clearly indicate that the exchange of the a3 domain within mouse class I antigens also influences the allodeterminants recog- nized by these T cell hybridomas.
88-3-24 34.1.2
173. 1
i B B
125 175.8
75 t 75 t
Q 58 100 158 2LlB 258 Q 58 iQ0 150 ZQQ 258
lA5. 1 L 141.0
Q 58 iQQ 158 208 250 Q 58 i B Q 15Q 288 258
i25 I 157.6 158.0 iQ0
.
25
I 0 5 0 i0Q 150 200 isQ Q 5Q 1013 150 260 250
Figure 3. Expression of Kb and KbKbDp hybrid antigens on L cells. FI are shown at the top of each figure. FI of the negative control (binding of anti-human class I1 mAb) was < 30.
1864 J.Yamamoto, M. Tanabe, M. Minami et al.
16.3
i
HTB 1 7 6 . 1 0
60
0 L
E m u - 4 0 0
Y u
2) 0
a N 2 0 2 I
s t i m u l a t o r c e l l s ( x 1 0 ” )
- 5
Q - = 4
H T B 1 7 7 . 2 o L K ~
a L K ~ K ~ D P / \ 0 LKb*
0 \
I - 10 1 0 9
s t i m u l a t o r c e l l s ( X I O - )
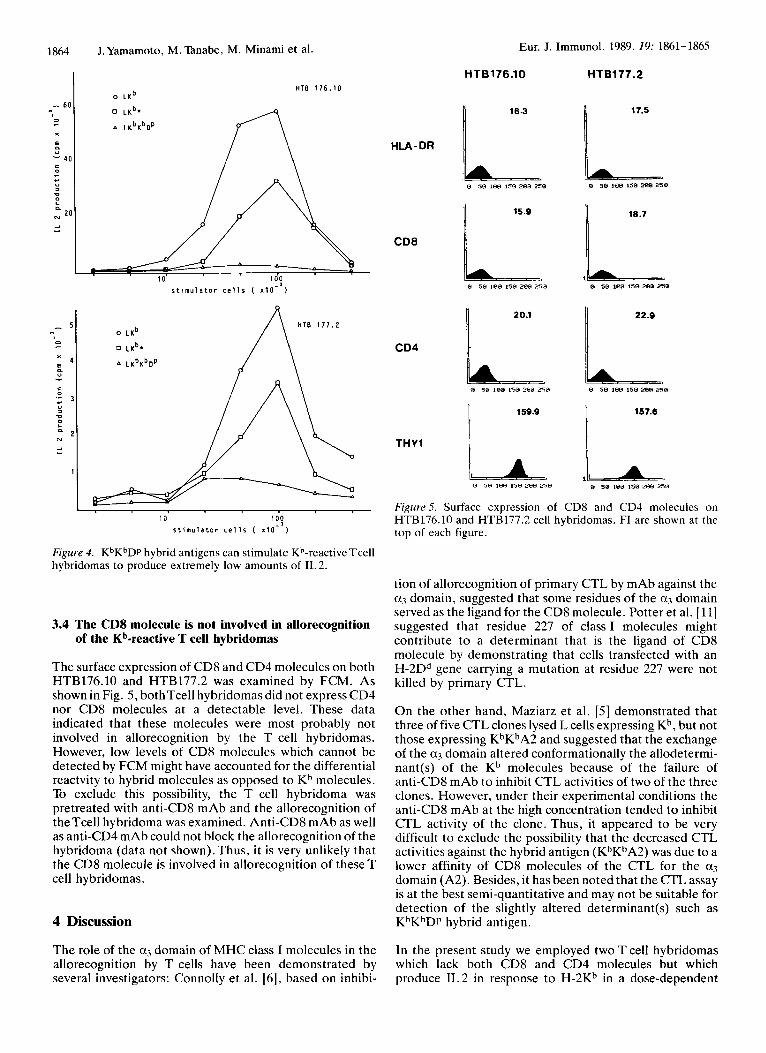
Figure 4. KbKbDp hybrid antigens can stimulate Kb-reactive Tcell hybridomas to produce extremely low amounts of IL 2.
3.4 The CD8 molecule is not involved in allorecognition of the Kb-reactive T cell hybridomas
The surface expression of CD8 and CD4 molecules on both HTB176.10 and HTB177.2 was examined by FCM. As shown in Fig. 5 , bothTcell hybridomas did not express CD4 nor CD8 molecules at a detectable level. These data indicated that these molecules were most probably not involved in allorecognition by the T cell hybridomas. However, low levels of CD8 molecules which cannot be detected by FCM might have accounted for the differential reactvity to hybrid molecules as opposed to Kh molecules. To exclude this possibility, the T cell hybridoma was pretreated with anti-CD8 mAb and the allorecognition of theTcell hybridoma was examined. Anti-CD8 mAb as well as anti-CD4 mAb could not block the allorecognition of the hybridoma (data not shown). Thus, it is very unlikely that the CD8 molecule is involved in allorecognition of these T cell hybridomas.
4 Discussion
The role of the a3 domain of MHC class I molecules in the allorecognition by T cells have been demonstrated by several investigators: Connolly et al. [6], based on inhibi-
HLA-DR
CD8
CD4
THY1
Eur. J. Immunol. 1989. 19: 1861-1865
HTB177.2 H T B 176.1 0
LA 0 SB I0H 150 ZBB 250
15.9 a 20.1 1 159.9 I
t
18.7
FigurrS. Surface expression of CD8 and CD4 molecules on HTB176.10 and HTB177.2 cell hybridomas. FI are shown at the top of each figure.
tion of allorecognition of primary CTL by mAb against the a3 domain, suggested that some residues of the a3 domain served as the ligand for the CD8 molecule. Potter et al. [ 111 suggested that residue 227 of class I molecules might contribute to a determinant that is the ligand of CD8 molecule by demonstrating that cells transfected with an H-2Dd gene carrying a mutation at residue 227 were not killed by primary CTL.
On the other hand, Maziarz et al. [5] demonstrated that three of five CTL clones lysed L cells expressing Kb, but not those expressing KbKbA2 and suggested that the exchange of the a3 domain altered conformationally the allodetermi- nant(s) of the Kb molecules because of the failure of anti-CD8 mAb to inhibit CTL activities of two of the three clones. However, under their experimental conditions the anti-CD8 mAb at the high concentration tended to inhibit CTL activity of the clone. Thus, it appeared to be very difficult to exclude the possibility that the decreased CTL activities against the hybrid antigen (KhKbA2) was due to a lower affinity of CD8 molecules of the CTL for the a3 domain (A2). Besides, it has been noted that the CTL assay is at the best semi-quantitative and may not be suitable for detection of the slightly altered determinant(s) such as KbKhDp hybrid antigen.
In the present study we employed two T cell hybridomas which lack both CD8 and CD4 molecules but which produce IL2 in response to H-2Kb in a dose-dependent

Figure 6. Amino acid sequence similarity between the a3 domains of
fashion. The expression of KbKbB7 and KbKbDp hybrid antigens on L cells in comparison with that of the parental Kb antigen were carefully determined and the levels of hybrid antigens were shown to be equivalent to those of parental Kb (LKb and LKb*), as evidenced by the very similar FI of these cells stained with four different anti- H-2Kb mAb. We were thus able to assess the response of T cell hybridomas via Tcell receptors to the hybrid antigens quantitatively in the absence of any influence of the accessory molecules.
Results of this study demonstrated that both T cell hybri- domas produce significantly less IL2 in response to KbKhB7 or KhKbDp than in response to Kb antigen. This would indicate that the a3 domain influences the structure of allodeterminant recognized by the Tcell hybridomas via Tcell receptors. Potter et al. [ll] reported that the single amino acid exchange at residue 227 in the a3 domain of H-2Dd antigen abrogated the reactivity of this antigen with primary CTL, but not with H-2Dd-reactive, IL 2-producing T cell hybridomas (3DT.52.5.8). From their FCM data, however, the expression level of Dd antigen of the mutant was higher than that of the native Dd molecules. This may account for the discrepancy between their results and ours, if theTcell hybridoma employed by these investigators also produced IL2 depending on the expression levels of Dd antigen on the surface of stimulator cells. As seen in Fig. 6, Kb and DP differ at 10 positions, while Kb and HLA-B7 differ at 25 positions. An amino acid unique to DP was observed at position 247, where Val in the Kb molecule was substituted by Leu in DP. Both amino acids are hydrophobic and very similar. Amino acids at the other nine positions are also present in other H-2 class I antigens. Thus, it is hard to believe that such a minimal difference in the sequence of the a3 domain creates a substantial conformational change in KbKbDp molecule and yet this subtle change in the intraspecies hybrid molecule was recognized by the Tcell hybridomas. X-ray crystallographic studies of HLA-A2 molecule [12] showed that several contacts between the a3 domain and the a1/ciz domains were observed at position 209,211,234 of the a3 domain and ten residues make contact with (32-microglobulin. The substitu- tions of amino acids at these positions in the a3 domain may cause a conformational change of allodeterminants of the a1 and a? domains. However, the amino acids in these positions are very conserved in both mouse and human class I molecules including H-2Kh. H-2Dp and HLA-B7 [13-161. This may suggest that the three-dimensional structure of Kb and DP presently under investigation might be slightly different from that of HLA-A2 molecule.
Several investigators [ 18, 191 recently demonstrated that other accessory molecules such as CD2 and LFA-1 onTcells were involved in allorecognition of T cells. However, it is well known that CD2 molecules bind LFA-3 molecules [20, 211 and LFA-1 molecules are a receptor for ICAM-1 [22, 23].We have no evidence that these molecules directly
MHC class I antigens.
bind class I molecules. Therefore, it is very unlikely that allorecognition of the chimeric molecules by these T cell hybridomas is influenced by the loss of binding of these accessory molecules on the epitope(s) of the a3 domain of class I molecule, although effect(s) of a binding loss of “unknown accessory molecules” still remains to be excluded. We thank Miss A i Kariyone f o r excellent technical assistance and Miss Mashu Noda f o r secretarial asristance. We also thank Dr. Jeffrey A. Frelingerfor helpfulsuggestions and reviewing the manu- script. Received March 30, 1989; in revised form June 27, 1989.
5 References
1 Allen, H.,Wraith, D., Para, P., Askonas, B. and Flavell, R. A , ,
2 Schaller, J., Shimonkewitz, R. , MacDonald, H. R. and Kvist,
3 Cowan, E. P., Jelachich, M. L., Coligan, J. E . and Biddison,W.
4 Layet, C., Kahn-Perles, B., Pontarotti, P., Ferrier, F!, Sire. J.
5 Maziarz, R. T.. Burakoff, S. J. and Bluestone, J. A,. J .
6 Connolly, J. M., Potter, T. T., Wormstall, E.-M. and Hansen,
7 Minami, M. , Kawasaki, H., Taira, S. and Nariuchi, H. ,
8 Minami, M., Nariuchi, H., Kawasaki, H. and Dorf, M.,
9 Tanabe, M., Takiguchi, M.,Yamamoto, J., Hayashi, H. and
Nature 1984. 309: 279.
S. , J. Exp. Med. 1986. 164: 1823.
E., Proc. Natl. Acad. Sci. U S A 1987. 84: 5014.
and Lemonnier, F. A., J. Immunol. 1987. 138: 2197.
Immunol. 1988. 140: 4372.
T. H. , J. Exp. Med. 1988. 168: 325.
J. Immunol. 1985. 135: 111.
J. Immunol. 1986. 136: 3341.
Kano, K., Transplantation 1989. 48: 135. 10 Ozato, K. and Sachs, D. H. , J . Immunol. 1981. 126: 317. 11 Potter, T. A., Bluestone, J. A. and Rajan, T. V., J. Exp. Med.
1988. 166: 956. 12 Bjorkman, F!,Saper, M. A. , Samraoui, B., Bennett, W. S..
Strominger, J. L. and Wiley, D. C., Nature 1987. 329: 506. 13 Schepart, B. S., Takahashi. H., Cozad, K. M., Morray, R. ,
Ozato, K.. Appela, E. and Frelinger, J. A.. J. Immunol. 1986. 136: 3489.
14 Weiss, E., Golden, L., Zakut, R., Mellor, A., Fahrner. K., Kvist, S. and Flavell, R. A,, E M B O J. 1983. 2: 453.
15 Kvist, S., Roverts, L. and Dobberstein, B., E M B O J. 1983. 2: 245.
16 Parham, P., Lomen. C. E., Lawlor, D. A. ,Ways, J. P., Holmes, N., Coppin, H. L.,Salter, R. D.,Wan, A. M. and Ennis. F! D.. Proc. Natl. Acad. Sci. USA 1988. 85: 4005.
17 Pottcr,T. A., Rajan,T.V., Dick, R. F., I1 and Bluestone. J. A.. Nature 1989. 337: 73.
18 Moingeon, P., Chang, H-C.. Wallner, B. P.. Stebbin, C., Frey, A. Z. and Reinherz, E. L., Nature 1989. 339: 312.
19 Altman, D. M., Hogg, N.,Trowsdale, J. and Wilkinson, D., Nature 1989. 338: 512.
20 Marlin, S. and Springer,T. A., Cell 1987. 51: 813. 21 Simmons. D.. Makgoba, M. and Seed, B., Nature 1989. 331:
22 Selvaraj, P.. Plunkett, M. L., Dustin, M., Sanders, M. E . , Shaw.
23 Shaw,S.. Lece, G. E. G., Quinones, R., Gress, R. E.. Springer,
624.
S. and Springer, T. A.. Nature 1987. 326: 400.
T. A. and Sanders, M. E., Nature 1986. 323: 262.


















