
Agrobacterium Protocols Volume 343 || Nicotiana (Nicotiana tobaccum, Nicotiana benthamiana)
12
143 From: Methods in Molecular Biology, vol. 343: Agrobacterium Protocols, 2/e, volume 1 Edited by: Kan Wang © Humana Press Inc., Totowa, NJ 12 Nicotiana (Nicotiana tobaccum, Nicotiana benthamiana) Tom Clemente Summary Agrobacterium-mediated transformation of Nicotiana species, namely, Nicotiana tobaccum and Nicotiana benthamiana, using leaf disks as the target explant has provided the plant community with a valuable tool for rapid evaluation of transgenes in higher plants. This protocol has a number of desirable attributes: readily available explant material, high efficiency, and a relatively quick turnaround time. The in vitro regenera- tion scheme of the leaf disks is prolific and follows an indirect organogenic differentia- tion. N. tobaccum and N. benthamiana are highly susceptible to a variety of disarmed A. tumefaciens strains, and the transformation system is amenable to a number of selec- tion agents. This protocol has proved to be easily transferable to a point at which a single investigator with minimal training can generate thousand of events per year. Moreover, seed size and quantity per event permit monitoring of segregation in Petri plates, and sufficient biomass can be accrued from an individual plant, which can be either clonally propagated or allowed to self-pollinate or easily outcrossed. Key Words: npt II; BAP; NAA; organogenesis; genetic engineering; Agrobacterium. 1. Introduction Nicotiana tobaccum and N. benthamiana have served as model species for the plant sciences for more than 20 yr. The latter possesses the unique trait of being susceptible to a vast number of plant viruses (> 500), even to those that are typically restricted to monocot hosts and, most surprisingly, even to several animal viruses (1). Consequently N. benthamiana has become a cornerstone for the study of host/virus interactions in plants. A criterion for a plant species to fall under the category of “model” is the ability to modulate gene expression rapidly and efficiently via plant genetic engineering. The seminal report describing a straightforward means to deliver transgenes into members of the Solanaceae family was communicated in the mid-1980s by Horsch et al. (2).
Transcript of Agrobacterium Protocols Volume 343 || Nicotiana (Nicotiana tobaccum, Nicotiana benthamiana)

Nicotiana 143
143
From: Methods in Molecular Biology, vol. 343: Agrobacterium Protocols, 2/e, volume 1Edited by: Kan Wang © Humana Press Inc., Totowa, NJ
12
Nicotiana (Nicotiana tobaccum, Nicotiana benthamiana)
Tom Clemente
SummaryAgrobacterium-mediated transformation of Nicotiana species, namely, Nicotiana
tobaccum and Nicotiana benthamiana, using leaf disks as the target explant has providedthe plant community with a valuable tool for rapid evaluation of transgenes in higherplants. This protocol has a number of desirable attributes: readily available explantmaterial, high efficiency, and a relatively quick turnaround time. The in vitro regenera-tion scheme of the leaf disks is prolific and follows an indirect organogenic differentia-tion. N. tobaccum and N. benthamiana are highly susceptible to a variety of disarmedA. tumefaciens strains, and the transformation system is amenable to a number of selec-tion agents. This protocol has proved to be easily transferable to a point at which a singleinvestigator with minimal training can generate thousand of events per year. Moreover,seed size and quantity per event permit monitoring of segregation in Petri plates, andsufficient biomass can be accrued from an individual plant, which can be either clonallypropagated or allowed to self-pollinate or easily outcrossed.
Key Words: npt II; BAP; NAA; organogenesis; genetic engineering; Agrobacterium.
1. IntroductionNicotiana tobaccum and N. benthamiana have served as model species for
the plant sciences for more than 20 yr. The latter possesses the unique trait ofbeing susceptible to a vast number of plant viruses (>500), even to those thatare typically restricted to monocot hosts and, most surprisingly, even to severalanimal viruses (1). Consequently N. benthamiana has become a cornerstonefor the study of host/virus interactions in plants. A criterion for a plant speciesto fall under the category of “model” is the ability to modulate gene expressionrapidly and efficiently via plant genetic engineering. The seminal reportdescribing a straightforward means to deliver transgenes into members of theSolanaceae family was communicated in the mid-1980s by Horsch et al. (2).

144 Clemente
This protocol has passed the test of time and remains a viable and efficientmeans to generate transgenic events from both N. tobaccum and N. benthamiana.Moreover, Agrobacterium-based gene delivery has advanced to where tran-sient gene expression in Nicotiana can be exploited to modulate gene expres-sion temporally in planta (3,4) thereby complementing the use of this powerfultool for functional genomics.
The Agrobacterium-mediated transformation protocol relies on an indirectorganogenic culture regime. This implies that differentiation of the explantfirst proceeds to a dedifferentiated callus phase prior to shoot organogenesis.The explant of choice is leaf disks isolated from 2–3-mo-old seedlings,although leaf disks obtained from older plants will suffice. Cell differentiationand transformation occurs about the parameter (cut edges) of the disks. Typi-cally, stable callus proliferation can be observed within 2 wk of selection andshoot organogenesis by 4 wk of culture. The derived shoots can be rootedwithin a 2-wk period and acclimated to the greenhouse. Total time from explantinoculation to establishing a primary transformant (T0) in the greenhouse is2 to 3 mo. The T0 plant will be hemizygous for the T-DNA. This time framemirrors that of the Arabidopsis floral dip method (5,6), but the Nicotiana pro-tocol necessitates additional labor and space requirements.
This transformation protocol is amenable to a number of selectable markergenes/selection agent combinations including but not limited to npt II/kanamy-cin (7), hpt/hygromycin (8), bar/glufosinate (9), and pmi/mannose (10). More-over, most genotypes on N. tobaccum are suitable; however, the workhorsegenotype for researchers has been ‘Xanthi’.
Plant genetic engineering protocols can be partitioned into two broadparameters, with the exception of the Arabidopsis floral dip method: (1) inte-gration of the foreign allele into a cell and (2) differentiation of that cell to alineage that will be transmitted to the next generation. The probability ofrecovering a functional transformant is equivalent to these two independentevents occurring. Hence, a plant species with high in vitro capacity that ishypersusceptible to A. tumefaciens will translate to high transformation effi-ciency, which is calculated on a transformant in soil per explant inoculatedbasis. This is assuming that the cells competent for in vitro morphogenesis arealso susceptible to A. tumefaciens infection. With respect to N. tobaccum andN. benthamiana, both can be classified as species with high in vitro capaci-tance and high susceptibility to A. tumefaciens; hence they possess a relativelyhigh transformation frequency. The ability to generate a large quantity oftransformants is desirable. However, the output potential of a plant geneticengineering protocol cannot compromise on quality, since the labor and spacecosts associated with screening poor-quality events may outweigh the benefitof a protocol with a high transformation frequency. The attributes, regenera-

Nicotiana 145
tion potential, and A. tumefaciens susceptibility translate to high output rates oftransgenic events with both Nicotiana species, but a significant portion of thederived events possess T-DNA insertions at multiple loci, which may lead to genesilencing (11). This potential problem can be alleviated by utilizing a less virulentstrain of A. tumefaciens such as the disarmed octopine strain LBA4404 (12).
The tobacco transformation frequency using the leaf disk protocol describedhere is relatively efficient. Genotypes of N. tobaccum will display a transfor-mation frequency ranging from 50 to 80%; frequencies with N. benthamianawill be significantly lower, approximately 20–30%.
2. Materials2.1. Plant Materials
1. Seed of N. benthamiana is indigenous to Australia. The species probably arosefrom a hybridization of two progenitors, N. suaveolens and N. debneyi.
2. Seed of N. tobaccum genotypes reported in the literature include SR-1, PetitHavana, and Xanthi. Although some genotype variation will exist in all geneticengineering protocols, most N. tobaccum genotypes are suitable for transformation.
2.2. Agrobacterium Strain and Constructs
1. A. tumefaciens strains: LBA4404/pAL4404 (12), C58C1/pPM90 (13), EHA101/pTiEHA101 (14), EHA105/pTiEHA105 (15), and NTL4/pTiEHA105 (16).
2. Binary vectors: selection of an appropriate binary vector will depend on theinherent antibiotic resistance of the selected A. tumefaciens strain (see Note 1).The small family of binary plasmids designated pPZP assembled by Hajdukiewiczet al. (17) constitute an excellent set of standard binary plasmids for use inAgrobacterium-mediated transformation. The backbone of the pPZP binary fam-ily also served as the foundation for the assembly of the well-distributedpCAMBIA vectors (www.cambia.org). In addition, Hellens et al. (18) wrote ashort guide to disarmed A. tumefaciens strains and commonly used binary plas-mids. This report summarizes the origin and antibiotic resistance of the respec-tive biological materials.
2.3. Stock Solutions
Major and minor stock solutions along with chelated iron stocks should bekept at 4°C. All growth regulator stocks made fresh monthly can be kept at4°C. Alternatively, they can be stored at –20°C. Prepared media plates can bestored at 4°C for up to 1 mo.
1. MS major salts (10X; 19), per liter: 19.0 g/L KNO3, 16.5 g/L NH4NO3, 3.7 g/LMgSO4·7H2O, 4.4 g/L CaCl2·2H2O, and 1.7 g/L KH2PO4 (see Note 2).
2. MS minor salts (100X; 19), per liter: 2.2 g/L MnSO4·4H2O, 83 mg/L KI, 620 mg/LH3BO4, 860 mg/L ZnSO4·7H2O, 2.5 mg/L CuSO4·5H2O, 25 mg/L NaMoO4·2H2O, and 2.5 mg/L CoCl2·6H2O.

146 Clemente
3. Iron stock (200X).
a. FeSO4·7H2O stock: dissolve 5.6 g of chemical in 1 L of dH2O. Keep theFeSO4·7H2O container wrapped with aluminum foil.
b. 0.5 M Na2EDTA stock (pH 8.0): to prepare, add 186.1 g of Na2EDTA to500 mL of distilled water. Monitor the pH while slowly adding pellets ofNAOH. The Na2EDTA will begin to dissolve as the pH approaches 8.0.Bring the final volume up to 1 L with distilled water.
Store the FeSO4·7H2O and Na2EDTA in separate containers at 4°C.
4. Vitamin B5 stock (100X; 20), per liter: 10 g/L myo-inositol, 100 mg/L nicotinicacid, 100 mg/L pryidoxine·HCl, and 1.0 g/L thiamine·HCl.
5. Growth regulator stocks (see Note 3).
a. Napthalene acetic acid (NAA): 1 mg/mL.b. Benzylaminopurine (BAP): 1 mg/mL.c. p-Chloro-phenoxy acetic acid (pCPA): 1 mg/mL.
6. Antibiotic stocks (see Note 4).
a. Carbenicillin: 50 mg/mL.b. Cefotaxime: 100 mg/mL.
2.4. Media
1. YEP medium (see Note 5): 10 g/L peptone, 5 g/L yeast extract, 5 g/L NaCl(pH 7.0). Sterilize by autoclaving. Add appropriate antibiotics when cooledto 55°C.
2. Preculture medium (1 L; see Note 6): 100 mL of major salt stock, 10 mL of minorsalt stock. 10 mL chelated iron prepared separately by mixing 5 mL of FeSO4
stock and 5 mL Na2EDTA stock and heat until solution turns yellow. Add 30 gof sucrose. Adjust pH of the medium with 1 N KOH to 5.7. Bring the medium upto 975 mL and add 8 g of grade A agar (Sigma). Sterilize by autoclaving.
Prepare vitamin and growth regulator solution. In 10 mL ddH2O, add 10 mLvitamin B5 stock, 1 mL BAP stock, 0.1 mL NAA stock, and 8 mg pCPA. Bringvitamin/growth regulator cocktail up to 25 mL with water and filter-sterilize.Add the sterilized vitamin/growth regulator cocktail to the autoclaved compo-nents once the temperature has reached 55°C.
3. Coculture medium (liquid 1 L; see Note 7): 10 mL of major salt stock, 1.0 mL ofminor salt stock, 10 mL chelated iron prepared separately by mixing 0.5 mL ofFeSO4 stock and 0.5 mL Na2EDTA stock and heat until solution turns yellow.Add 30 g of sucrose and 3.7 g 2-morpholinoethanesulfonic acid (MES) buffer(pH 5.4). Bring the medium up to 950 mL.
Prepare vitamin and growth regulator solution. In 10 mL of ddH2O, add10 mL vitamin stock, 1 mL BAP stock, 0.1 mL NAA stock, and 8 mg of pCPA.Dissolve 38 mg of acetosyringone. Bring vitamin/growth regulator cocktail upto 50 mL with water and filter-sterilize. Add the sterilized vitamin/growth

Nicotiana 147
regulator cocktail to the autoclaved components once the temperature hasreached 55°C.
4. Selection medium (1 L; see Note 8): 100 mL of major salt stock, 10 mL of minorsalt stock, 10 mL chelated iron prepared separately by mixing 5 mL of FeSO4
stock and 5 mL Na2EDTA stock and heat until solution turns yellow. Add 30 gof sucrose. Adjust pH of the medium with 1 N KOH to 5.7. Bring the medium upto 975 mL and add 8 g of grade A agar (Sigma). Sterilize by autoclaving.
Prepare vitamin and growth regulator solution. In 10 mL ddH2O add 10 mLvitamin B5 stock, 1 mL BAP stock, 0.1 mL NAA stock. Add 2 mL carbenicillinstock and 1 mL of cefotaxime stock. Bring vitamin/growth regulator cocktail upto 25 mL with water and filter-sterilize. If using nptII add 150 mg/L kanamycin.If using hpt add 10 mg/L hygromycin. If using bar add 3 mg/L glufosinate.Add the sterilized vitamin/growth regulator cocktail to the autoclaved compo-nents once the temperature has reached 55°C.
5. Rooting medium (1 L; see Note 9): 50 mL of major salt stock, 5 mL of minor saltstock, 10 mL chelated iron prepared separately by mixing 5 mL FeSO4 stock and5 mL Na2EDTA stock and heat until solution turns yellow. Add 10 g sucrose.Adjust pH of the medium with 1 N KOH to 5.7. Bring the medium up to 975 mLand add 8 g of grade A agar (Sigma). Sterilize by autoclaving.
Prepare vitamin and growth regulator solution. In 10 mL ddH2O add, 10 mLvitamin B5 stock, 0.1 mL of NAA stock. Add 2 mL carbenicillin stock and 1 mLof cefotaxime stock. Bring vitamin/growth regulator cocktail up to 25 mL withwater and filter-sterilize. If using nptII add 75 mg/L kanamycin. If using hpt add5 mg/L hygromycin. If using bar add 3 mg/L glufosinate. Add the sterilized vita-min/growth regulator cocktail to the autoclaved components once the tempera-ture has reached 55°C
6. Germination medium (1 L): 100 mL of major salt stock, 10 mL of minor saltstock, 10 mL chelated iron prepared separately by mixing 5 mL FeSO4 stock and5 mL Na2EDTA stock and heat until solution turns yellow. Add 30 g of sucrose.Adjust pH of the medium with 1 N KOH to 5.7. Bring the medium up to 975 mLand add 8 g of grade A agar (Sigma). Sterilize by autoclaving.
Prepare vitamin solution by taking 10 mL of vitamin B5 stock and bringing itup to 25 mL with water. Filter-sterilize and add to the autoclaved componentsonce the temperature has reached 55°C.
2.5. Other Supplies
1. Sundae cup vessels: Sweetheart™ lids, cat. no. LDS58: base, cat. no. DSD8X.2. Jiffy™-cup: model 122 (Hummert™ International, Earth City, MO).3. Soil: Metro-Mix 360 (Hummert International).4. Magenta™ vessel: cat. no. V8505 (Sigma).5. Hoagland’s solution: cat. no. H2395 (Sigma).6. Whatman filter paper: cat. no. 1001070 (VWR).

148 Clemente
3. Methods3.1. Plant Material Preparation (see Note 10)
3.1.1. From Greenhouse-Grown Plants1. Select 4- to 6-inch (N. benthamiana leaves are typically smaller than N. tobaccum)
leaves from 1- to 2-mo-old N. benthamiana and N. tobaccum plants growingunder greenhouse conditions. The upper two fully expanded leaves are sufficientfor 30 to 40 leaf disks.
2. Place the selected leaves in a large beaker and cover with distilled water for 20 to30 min.
3. Drain the water and cover the leaves with 10% commercial bleach (0.6% sodiumhypochlorite) supplemented with 1 drop of Tween-20 per 50 mL. Allow the leavesto soak in the bleach solution for 10 min (see Note 11). Rinse five times withsterile distilled water.
3.1.2. From Seeds1. Place approximately 50 L packed volume of seeds from the respective species
in an Eppendorf tube, sterilized by exposure to 1 mL of 100% ethanol for 1 min.2. Following the ethanol wash, briefly centrifuge for at 16,000g for 15 s.3. Remove ethanol and replace with 1 mL 50% commercial bleach (3% sodium
hypochlorite) supplemented with Tween-20 (1 drop per 50 mL). Vortex briefly.Let the seed sit in the bleach solution for 15 min.
4. Centrifuge the seed at 16,000g for 30 s and draw off bleach with a pipet.5. Wash the sterilized seed with sterile distilled water five times by repeating step 4.6. Transfer the sterilized seeds to a 15-mL Falcon tube. Add 8 mL of 0.4% water
agar supplemented with 100 mg/L cefotaxime. Mix the seed/water agar well.7. Evenly overlay the mixture on top of the germination medium poured in sundae
cup vessels. Allow the seedlings to grow for up to 1 mo, under an 18-h lightregime (approximate light intensity of 140 mol/m2/s) at 24–28°C.Explants prepared from in vitro germinated seeds will not require the steriliza-tion procedure outlined in Subheading 3.1.1.
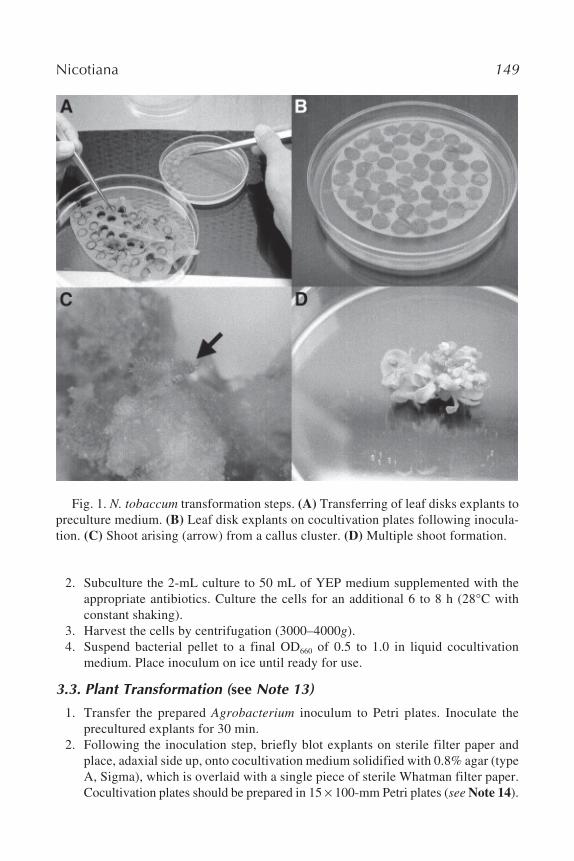
3.1.3. Leaf Explant Preparation1. Stack prepared leaves in a Petri plate. Prepare disks using a sterile cork borer (5–
8 mm). Avoid leaf margins and mid rib (Fig. 1A).2. Place explants adaxial side up onto the preculture medium (use 15 100-mm
Petri plates). Preculture approximately 40 explants per plate. Culture the explantsat 25°C under an 18-h light regime (approximate light intensity of 140 mol/m2/s)for 24 h.
3.2. Preparation of Agrobacterium Culture (see Note 12)
1. Initiate a 2-mL culture of A. tumefaciens in YEP medium supplemented with theappropriate antibiotics. Culture the cells at 28°C with constant shaking. Allowthe culture to become saturated. Saturation usually requires 2 to 4 h of culture.
Nicotiana 149
2. Subculture the 2-mL culture to 50 mL of YEP medium supplemented with theappropriate antibiotics. Culture the cells for an additional 6 to 8 h (28°C withconstant shaking).
3. Harvest the cells by centrifugation (3000–4000g).4. Suspend bacterial pellet to a final OD660 of 0.5 to 1.0 in liquid cocultivation
medium. Place inoculum on ice until ready for use.
3.3. Plant Transformation (see Note 13)
1. Transfer the prepared Agrobacterium inoculum to Petri plates. Inoculate theprecultured explants for 30 min.
2. Following the inoculation step, briefly blot explants on sterile filter paper andplace, adaxial side up, onto cocultivation medium solidified with 0.8% agar (typeA, Sigma), which is overlaid with a single piece of sterile Whatman filter paper.Cocultivation plates should be prepared in 15 100-mm Petri plates (see Note 14).
Fig. 1. N. tobaccum transformation steps. (A) Transferring of leaf disks explants topreculture medium. (B) Leaf disk explants on cocultivation plates following inocula-tion. (C) Shoot arising (arrow) from a callus cluster. (D) Multiple shoot formation.

150 Clemente
3. Cocultivate the explants at 24°C, 18-h light regime for 3 d. Cocultivate approxi-mately 40 explants per plate (Fig. 1B).
4. Following the 3-d cocultivation period, transfer the explants to regenerationmedium supplemented with the appropriate selection agent. Regenerationmedium should be prepared in 15 100-mm Petri plates. Culture up to 10 explantsper plate and incubate the plates at 28°C under an 18-h light regime. Transfer theexplants to fresh regeneration medium every 2 wk.
5. Callus clusters should be apparent forming along the cut parameter of the diskbeginning after 2 wk of selection. Multiple shoots can arise from a single calluscluster, and multiple clusters can form from a single disk. Shoots arising from asingle cluster will generally be clones. An example of a shoot arising from acallus cluster is shown in Fig. 1C; multiple shoot proliferation from a callus clus-ter is displayed in Fig. 1D. Therefore, be sure to track the shoots arising from thesame cluster.
6. Excise shoots individually once they reach a length greater than 3 mm and trans-fer to rooting medium (see Note 15) supplemented with the appropriate selectionagent. Rooting medium should be poured in a sundae cup vessel or equivalent toallow for sufficient plantlet development.
7. The rooting stage is carried out under the same environmental conditions usedduring the regeneration step. Roots should be adequately established within2 wk. A putative N. tobaccum primary transformant ready for acclimation to soilis shown in Fig. 2.
Fig. 2. N. tobaccum primary transformant rooted and ready for acclimation step.

Nicotiana 151
3.4. Plant Care and Seed Harvest (see Note 16)
1. Carefully remove plantlets with a well-established root system from rootingmedium. Wash excess agar off the root system with tap water, transfer the plant-let to a Jiffy-cup, and cover with Metro-Mix 360.
2. Place the Jiffy-cup into a double stacked Magenta vessel and water as neededwith 1/2X Hoagland’s solution.
3. Place the Magenta vessel into a growth chamber under an 18-h light regime andtemperature of 28°C.
4. After 2 to 3 d crack the lid of the stacked Magenta vessel. Once the plantlet isfully acclimated, transfer to the greenhouse by transplanting the Jiffy-cup to a6-inch pot and cover containing standard greenhouse soil mix.
3.5. Progeny Test and Seed Storage
1. T1 seed should be collected from fully mature pods. Seeds can be stored in coinenvelopes or Eppendorf tubes. Ideal storage temperature for seed is 8°C underlow humidity; however, seed viability at room temperature or 4°C remains highfor a number of years.
2. To conduct segregation analysis, sterilize harvested T1 seed, using the steriliza-tion technique described in Subheading 3.1.2.
3. After the last washing, transfer the seed to a 15-mL Falcon tube. Add 8 mL of0.4% water agar supplemented with 100 mg/L cefotaxime. Mix the water agar/seed mixture well by inverting the Falcon tube 5 to 10 times.
4. Overlay agar/seed mixture onto a 100 150-mm Petri plate containing approxi-mately 45 mL of germination medium supplemented with the appropriate selec-tion agent. Allow the plates to air-dry with the lids off for approximately 10 minunder a laminar flow hood.
5. Place the plates at 28°C under an 18-h light regime. Take segregation data fol-lowing 2 wk of germination.
4. Notes1. To select antibiotic compatibility between the A. tumefaciens strain and binary
plasmid, it is necessary to know the respective drug resistance of the biologicalmaterial. For example, A. tumefaciens strain C58C1 carrying the disarmedTi-plasmid pPMP90 (13) is resistant to rifampicin (chromosome marker) at atypical use rate of 50 mg/L and gentamicin (Ti-plasmid drug resistance).
2. Store the major and minor salt stock solutions at 4°C.3. NAA dissolves readily in water. To prepare BAP stock, dissolve powder in
1 M NAOH and bring up in distilled water. pCPA is prepared fresh in 95% ethanol.4. The antibiotic stocks of carbenicillin and cefotaxime are prepared in water.
The stocks may be stored at 4°C for 30 d or frozen in aliquots at –20°C forprolonged storage.
5. Be familiar with the inherent marker genes in the A. tumefaciens strains beingused and those of the corresponding binary plasmid (see Note 1).

152 Clemente
6. Pour preculture medium in 15 100-mm Petri plates. Plates can be stored for upto 1 mo at 4°C. After chelated iron has been separately prepared, add it directly tothe medium prior to autoclaving. Allow the medium to cool in a water bath set at55°C prior to adding vitamin and growth regulator cocktail solution.
7. Liquid coculture medium can be stored at 4°C for 1 mo. Acetosyringone shouldbe dissolved with a few drops of DMSO first, prior to adding to the vitamin/growth regulator cocktail solution. Pour solid coculture medium in 15 100-mmPetri plates. Plates can be stored for up to 1 mo at 4°C.
Cocultivation medium is designed to optimize induction of A. tumefaciens,without compromising tobacco cell culture. Hence, reduced salts, low pH, andthe inducing agent acetosyringone will provide conditions to maximize for virgene induction with minimal compromise to the in vitro culture of the explant.Preventing A. tumefaciens cells from engulfing the explants during the cocultivationstep is aided by filter paper overlays on the cocultivation plates. This in turn mayreduce browning of the explants.
8. The selection medium is poured in 15 100-mm Petri plates. Antibiotic stocksfor kanamycin and hygromycin should be prepared in water and the concentra-tion based on active ingredients, not merely mass.
9. Salts and sucrose are reduced during the rooting phase. This will tend to increaseroot induction frequencies.
10. If a series of tobacco transformations will be required, a constant supply ofexplant material can be established by biweekly seeding flats of tobacco. Culturethe stock plants under greenhouse conditions using standard soil mix or Metro-mix 360. The seedlings can be maintained in flats for 2–3 mo.
11. Care must be taken not to soak the leaf tissue in the bleach solution for too long.This will depend on the age of the leaf selected. Generally 8 to 10 min is sufficient.
12. For long-term storage A. tumefaciens strains should be kept at –80°C in 25%glycerol. Streak the A. tumefaciens strain selected for transformation on a YEPplate supplemented with the appropriate antibiotics and culture at 28°C. It willtake about 48 h for the culture to grow sufficiently from a glycerol stock.The plate culture should be used to initiate the liquid inoculum. Starting from aplate will help ensure a clean culture.
13. Transfer the leaf disks from the preculture plate directly to the A. tumefaciensinoculum. Be sure the explants are not merely floating on top of the inoculum.Inoculate the explants for 30 min, agitating the bacterium once every 10 min.
14. At the end of the inoculation step, immediately transfer the explants to thecocultivation plates. Following the cocultivation period, transfer the explants tothe regeneration medium. Here reduce the explant density per plate to 10.After 1 wk on regeneration plates, check the explants for any visible sign ofcontamination. If a contaminated explant is observed on a plate, transfer all otherexplants to a fresh plate and discard the contaminants.
15. Do not try and root more than four shoots per sundae cup. Sweetheart sundaecups per se are not labeled “guaranteed sterile,” but if the sleeves are handled likestacks of Petri plates, contamination associated with use of these vessels should

Nicotiana 153
not be a problem. Once roots have been initiated, the plantlets are ready to beestablished in soil. There is no need to wait for a massive root system to develop.
16. Once a plantlet is transferred out of culture, it should be identified with a uniqueevent number. Keep the plantlet in the stacked Magenta vessel for about 2 to 3 d.After this period crack the upper Magenta so that it sits on the connecter withaeration around the corners. If no signs of stress occur on the plantlet after2 additional days with the upper lid ajar, then the event is ready to be establishedin the greenhouse. Both N. benthamiana and N. tobaccum are relatively hardy.Establishing primary transformants out of culture is rarely problematic.
References1. Dasgupta, R., Garcia, B. H. 2nd, and Goodman, R. M. (2001) Systemic spread of
an RNA insect virus in plants expressing plant viral movement protein genes.Proc. Natl. Acad. Sci. USA 98, 4910–4915.
2. Horsch, R. B., Fry, J. E., Hoffmann, N. L., Eichholtz, D., Rogers, S. G., and Fraley,R. T. (1985) A simple and general method for transferring genes into plants.Science 227, 1229–1231.
3. Voinnet, O., Rivas, S., Mestre, P., and Baulcombe, D. (2003) An enhanced tran-sient expression system in plants based on suppression of gene silencing by thep19 protein of tomato bushy stunt virus. Plant J. 33, 949–956.
4. Bendahmane, A., Querci, M., Kanyuka, K., and Baulcombe, D. C. (2000)Agrobacterium transient expression system as a tool for the isolation of diseaseresistance genes: application to the Rx2 locus in potato. Plant J. 21, 73–81.
5. Bechtold, N., Ellis, J., and Pelletier, G. (1993) In planta Agrobacterium mediatedgene transfer by infiltration of adult Arabidopsis thaliana plants. C. R. Acad. Sci.Paris 316, 1194–1199.
6. Clough, S. J. and Bent, A. F. (1998) Floral dip: a simplified method forAgrobacterium-mediated transformation of Arabidopsis thaliana. Plant J. 16,735–743.
7. Fraley, R. T., Rogers, S. G., Horsch, R. B., et al. (1983) Expression of bacterialgenes in plant cells. Proc. Natl. Acad. Sci. USA 80, 4803–4807.
8. Gritz, L. and Davies, J. (1983) Plasmid-encoded hygromycin B resistance:the sequence of hygromycin B phosphotransferase gene and its expression inEscherichia coli and Saccaromyces cerevisiae. Gene 26, 179–188.
9. Thompson, C. J., Movva, N. R., Tizard, R., et al. (1987) Characterization of theherbicide-resistance gene bar from Streptomyces hygroscopicus. EMBO J. 6,2519–2523.
10. Joersbo, M. and Okkels, F. T. (1996) A novel principle for selection of transgenicplant cells: positive selection. Plant Cell Rep. 16, 219–221.
11. Cerutti, H. (2003) RNA interference: traveling in the cell and gaining functions?Trends Genet. 19, 39–46.
12. Hoekema, A., Hirsch, P. R., Hooykaas, P. J. J., and Schilperoort, R. (1983)A binary plant vector strategy based on separation of vir- and T-region of theAgrobacterium tumefaciens Ti-plasmid. Nature 303, 179–180.

154 Clemente
13. Koncz, C. and Schell, J. (1986) The promoter of TL-DNA gene 5 controls thetissue-specific expression of chimaeric genes carried by a novel type of Agro-bacterium binary vector. Mol. Gen. Genet. 204, 383–396.
14. Hood, E. E., Helmer, G. L., Fraley, R. T., and Chilton, M.-D. (1986)The hypervirulence of Agrobacterium tumefaciens A281 is encoded in a regionof pTiBo542 outside of T-DNA. J. Bacteriol. 168, 1291–1301.
15. Hood, E. E., Gelvin, S. B., Melchers, L. S., and Hoekema, A. (1993) New Agro-bacterium helper plasmids for gene transfer to plants. Transgenic Res. 2, 208–218.
16. Luo, Z.-Q., Clemente, T., and Farrand, S. K. (2001) Construction of a derivativeof Agrobacterium tumefaciens C58 that does not mutate to tetracycline resistance.Mol. Plant Microbiol. Int. 14, 98–103.
17. Hajdukiewicz, P., Svab, Z., and Maliga, P. (1994) The small, versatile pPZP fam-ily of Agrobacterium binary vectors for plant transformation. Plant Mol. Biol. 25,989–994.
18. Hellens, R., Mullineaux, P., and Klee, H. (2000) Technical focus: a guide toAgrobacterium binary Ti vectors. Trends Plant Sci. 5, 446–451.
19. Murashige, T. and Skoog, F. (1962) A revised medium for rapid growth and bio-assays with tobacco tissue cultures. Physiol. Plant 15, 473–497.
20. Gamborg, O. L., Miller, R. A., and Ojima, K. (1968) Nutrient requirements ofsuspension cultures of soybean root cells. Exp. Cell Res. 50, 152–158.





![Lib Tesis Revistas ibros - CICY.mxde TRXh1, NPR1 y PR1 en Nicotiana benthamiana en una infección mixta de begomovirus. Mérida, Yuc.. xi, 93 p. . [TM A583 2014] (1 ejemplar) Avila](https://static.fdocument.pub/doc/165x107/5f6fb340b1979211ac3f2636/lib-tesis-revistas-ibros-cicymx-de-trxh1-npr1-y-pr1-en-nicotiana-benthamiana.jpg)
![Transgenik Melalui Crown Gall (Melalui Agrobacterium Tumefaciens)[1]](https://static.fdocument.pub/doc/165x107/55cf900b550346703ba2a64a/transgenik-melalui-crown-gall-melalui-agrobacterium-tumefaciens1.jpg)











